Das Uracil Es ist eine stickstoffhaltige Base vom Pyrimidin-Typ, die in Ribonukleinsäure (RNA) vorkommt. Dies ist eine der Eigenschaften, die RNA von Desoxyribonukleinsäure (DNA) unterscheiden, da letztere Thymin anstelle von Uracil enthält. Beide Substanzen, Uracil und Thymin, unterscheiden sich nur darin, dass die zweite eine Methylgruppe aufweist.
Aus evolutionärer Sicht wurde vorgeschlagen, dass RNA das erste Molekül war, das genetische Informationen speicherte und als Katalysator in Zellen vor DNA und Enzymen fungierte. Aus diesem Grund wird angenommen, dass Uracil eine Schlüsselrolle in der Entwicklung des Lebens gespielt hat.
In Lebewesen kommt Uracil nicht in freier Form vor, sondern bildet üblicherweise die Nukleotide Monophosphat (UMP), Diphosphat (UDP) und Triphosphat (UTP). Diese Uracil-Nukleotide haben unterschiedliche Funktionen, wie die RNA- und Glykogen-Biosynthese, die isomere Umwandlung von Zuckern und die Regulation der Glutaminsynthase..
Artikelverzeichnis
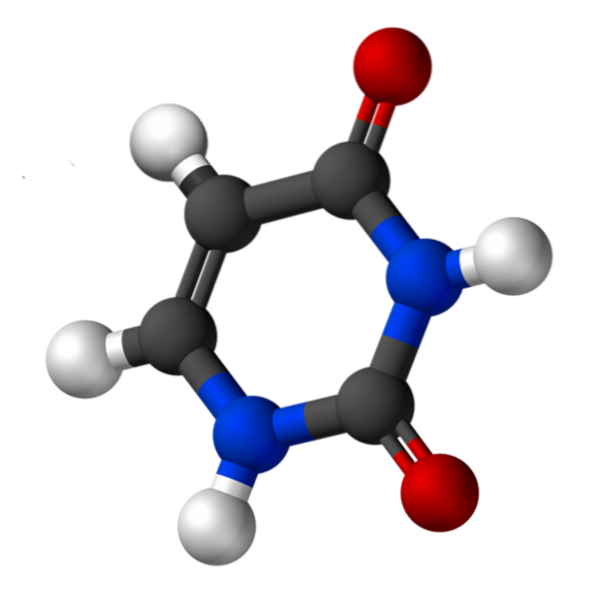
Uracil, genannt 2,4-Dioxypyridin, hat die empirische Formel C.4H.4N.zweiODERzwei, dessen Molekulargewicht 112,09 g / mol beträgt und als weißes Pulver gereinigt wird.
Die Struktur von Uridin ist ein heterocyclischer Ring mit vier Kohlenstoffatomen und zwei Stickstoffatomen mit alternierenden Doppelbindungen. Ist planar.
Es hat eine Löslichkeit von 50 mg / ml bei 25 ° C in 1 M Natriumhydroxid und einen pKa zwischen 7,9 und 8,2. Die Wellenlänge, bei der die maximale Absorption auftritt (ʎmax) liegt zwischen 258 und 260 nm.
Es gibt einen gemeinsamen Weg für die Pyrimidinnukleotidbiosynthese (Uracil und Cytokin). Der erste Schritt ist die Biosynthese von Carbamoylphosphat aus COzwei und NH4+, welches durch Carbamoylphosphatsynthetase katalysiert wird.
Pyrimidin wird aus Glasphosphat und Aspartat hergestellt. Beide Substanzen reagieren und bilden N-Carbamoylaspartat, eine Reaktion, die durch Aspartat-Transcabamoylase (ATCase) katalysiert wird. Der Verschluss des Pyrimidinrings wird durch durch Dihydrootase katalysierte Dehydratisierung verursacht und erzeugt L-Dihydrorotat..
L-Dihydrorotat wird oxidiert und in Orotat umgewandelt; Elektronenakzeptor ist NAD+. Es ist eine durch Dihydroorotatdehydrogenase katalysierte Reaktion. Der nächste Schritt ist die Übertragung der Phosphoribosylgruppe von Phosphoribosylpyrophosphat (PRPP) auf Orotat. Bildet Orotidylat (OMP) und anorganisches Pyrophosphat (PPi), katalysiert durch Orotatphosphoribosyltransferase.
Der letzte Schritt besteht in der Decarboxylierung des Pyrimidinrings des Orotidylats (OMP). Bildet Uridylat (Uridin-5'-monophosphat, UMP), das durch eine Decarboxylase katalysiert wird.
Dann wird durch die Beteiligung einer Kinase eine Phosphatgruppe von ATP auf UMP übertragen, wobei UDP (Uridin-5'-diphosphat) gebildet wird. Letzteres wird wiederholt, wobei UTP (Uridin-5'-triphosphat) gebildet wird..
In Bakterien erfolgt die Regulation der Pyrimidin-Biosynthese durch negative Rückkopplung auf der Ebene der Aspartat-Transcabamoylase (ATCase)..
Dieses Enzym wird durch CTP (Cytidin-5'-triphosphat) gehemmt, das das Endprodukt des Pyrimidin-Biosynthesewegs ist. ATCase hat regulatorische Untereinheiten, die an den allosterischen Regulator CTP binden.
Bei Tieren erfolgt die Regulation der Pyrimidin-Biosynthese durch negative Rückkopplung auf der Ebene von zwei Enzymen: 1) Carbamoylphosphatsynthase II, die durch UTP inhibiert und durch ATP und PRPP aktiviert wird; und 2) OMP-Decarboxylase, die durch das Produkt der von ihr katalysierten Reaktion, UMP, inhibiert wird. Die Geschwindigkeit der Biosynthese von OMP variiert mit der Verfügbarkeit von PRPP.
Uracil ist in allen Arten von RNA vorhanden, wie Messenger-RNA (mRNA), Transfer-RNA (tRNA) und ribosomaler RNA (rRNA). Die Biosynthese dieser Moleküle erfolgt durch einen Prozess, der als Transkription bezeichnet wird..
Während der Transkription wird die in der DNA enthaltene Information durch eine RNA-Polymerase in RNA kopiert. Der umgekehrte Prozess, bei dem die in der RNA enthaltenen Informationen in DNA kopiert werden, findet bei einigen Viren und Pflanzen durch reverse Transkriptase statt..
Die RNA-Biosynthese erfordert Nucleosidtriphosphat (NTP), nämlich: Uridintriphosphat (UTP), Cytidintriphosphat (CTP), Adenintriphosphat (ATP) und Guanintriphosphat (GTP). Die Reaktion ist:
(RNA)n Rückstände + NTP -> (RNA)n + 1 Rückstand + PPi
Die Hydrolyse von anorganischem Pyrophosphat (PPi) liefert die Energie für die RNA-Biosynthese.
Zuckerester sind in lebenden Organismen sehr verbreitet. Einige dieser Ester sind Nukleosidester-Diphosphate wie UDP-Zucker, die in Zellen sehr häufig vorkommen. UDP-Zucker sind an der Biosynthese von Disacchariden, Oligosacchariden und Polysacchariden beteiligt.
In Pflanzen erfolgt die Saccharosebiosynthese über zwei Wege: einen primären und einen sekundären.
Der Hauptweg ist der Transfer von D-Glucose von UDP-D-Glucose zu D-Fructose unter Bildung von Saccharose und UDP. Der sekundäre Weg umfasst zwei Schritte: Er beginnt mit UDP-D-Glucose und Fructose-6-phosphat und endet mit der Bildung von Saccharose und Phosphat..
In den Brustdrüsen erfolgt die Laktosebiosynthese aus UDP-D-Galaktose und Glukose.
In Pflanzen wird die Cellulosebiosynthese durch kontinuierliche Kondensation von Beta-D-Glucosylresten von UDP-Glucose bis zum nichtreduzierenden Ende der wachsenden Polyglucosekette durchgeführt. In ähnlicher Weise erfordert die Amylose- und Amylopektin-Biosynthese UDP-Glucose als Glucosespendersubstrat für die wachsende Kette..
Bei Tieren werden sowohl UDP-Glucose als auch ADP-Glucose für die Glykogenbiosynthese verwendet. In ähnlicher Weise erfordert die Chondroitinsulfat-Biosynthese UDP-Xylose, UDP-Galactose und UDP-Glucuronat..
Die Umwandlung von Galactose in ein Glykolyse-Zwischenprodukt erfolgt über den Leloir-Weg. Einer der Schritte auf diesem Weg wird durch das Enzym UDP-Galactose-4-Epimerase katalysiert, das die Umwandlung von UDP-Galactose in UDP-Glucose erleichtert..
Während der Glykoprotein-Biosynthese kreuzen Proteine die cis-, mittleren und trans-Säcke des Golgi-Apparats.
Jeder dieser Säcke enthält eine Reihe von Enzymen, die Glykoproteine verarbeiten. Zuckermonomere wie Glucose und Galactose werden dem Protein Oligosaccharid aus UDP-Hexose und anderen Nucleotiden-Hexose zugesetzt.
Die Hexose-Nukleotide werden per Antiport zu den Golgi-Zisternen transportiert. UDP-Galactose (UDP-Gal) und UDP-N-Acetylgalactosamin (UDP-GalNAc) gelangen durch Austausch gegen UMP aus dem Cytosol in die Zisternen.
In der Golgi-Zisterne hydrolysiert eine Phosphatase eine Phosphatgruppe an UDP und bildet UMP und Pi. UDP stammt aus Reaktionen, die durch Galactosyltransferase und N-Acetylgalactosamyltransferase katalysiert werden. Durch Phosphatase gebildetes UMP dient zum Nucleotid-Hexose-Austausch.
Ein Regulationsmechanismus der Glutaminsynthase ist die kovalente Modifikation, die aus einer Adenylierung besteht, die sie inaktiviert, und einer Dedenylierung, die sie aktiviert. Diese kovalente Modifikation ist reversibel und wird durch Adenyltransferase katalysiert..
Die Adenyltransferaseaktivität wird durch die Bindung des PII-Proteins moduliert, das durch eine kovalente Modifikation, die Uridinylierung, reguliert wird.
Sowohl die Uridylierung als auch die Deuridylierung werden durch Uridylyltransferase durchgeführt. In diesem Enzym beruht die Uridylierungsaktivität auf Glutamin und Phosphat und wird durch die Bindung von Alpha-Ketoglutarat und ATP an PII aktiviert.
Einige mRNAs werden vor der Translation bearbeitet. In einigen eukaryotischen Organismen wie Trypanosoma brucei, Es gibt eine RNA-Bearbeitung des Gentranskripts der Cytochromoxidase-Untereinheit II. Dies geschieht durch die Insertion von Uracilresten, eine Reaktion, die durch die terminale Uridyltransferase katalysiert wird..
Eine zum bearbeiteten Produkt komplementäre Leit-RNA dient als Vorlage für den Bearbeitungsprozess. Die zwischen dem anfänglichen Transkript und der Leit-RNA gebildeten Basenpaare implizieren G = U-Basenpaare, die nicht Watson-Crick sind und in der RNA häufig vorkommen..
Unter physiologischen Bedingungen ist die Biosynthese von Glykogen aus Glucose-1-phosphat thermodynamisch unmöglich (ΔG positiv). Aus diesem Grund erfolgt vor der Biosynthese die Aktivierung von Glucose-1-phosphat (G1P). Diese Reaktion kombiniert G1P und UTP, um Uridindiphosphatglucose (UDP-Glucose oder UDPG) zu bilden..
Die Reaktion wird durch UDP-Glucosepyrophosphorylase katalysiert und ist wie folgt:
G1P + UTP -> UDP-Glucose + 2Pi.
Die Variation der freien Gibbs-Energie in diesem Schritt ist groß und negativ (-33,5 KJ / mol). Während der Reaktion auf Sauerstoff greift G1P das Alpha-Phosphoratom von UTP an und bildet UDP-Glucose und anorganisches Pyrophosphat (PPi). Als nächstes wird das PPi durch eine anorganische Pyrophosphatase hydrolysiert, deren Hydrolyseenergie die allgemeine Reaktion antreibt.
UDP-Glucose ist eine "energiereiche" Substanz. Es ermöglicht die Bildung der glykosidischen Bindungen zwischen dem Glucoserest und der wachsenden Polysaccharidkette. Das gleiche energetische Prinzip gilt für Reaktionen, an denen UDP-Zucker beteiligt sind, wie die Biosynthese von Disacchariden, Oligosacchariden und Glykoproteinen..
Es gibt DNA-Läsionen, die spontan auftreten. Eine dieser Läsionen ist die spontane Desaminierung von Zytokin und die daraus resultierende Umwandlung in Uracil. In diesem Fall erfolgt die Reparatur durch Entfernen der modifizierten DNA-Base durch ein Enzym namens Uracil-DNA-Glycosylase..
Das Enzym Uracil-DNA-Glycosylase entfernt das beschädigte Cytokin (Uracil) und erzeugt einen Desoxyribose-Rest, dem die Stickstoffbase fehlt, die als AP-Stelle (Apurin-Apyrimidin-Stelle) bezeichnet wird..
Das Enzym AP-Endonuklease schneidet dann das Phosphodiester-Rückgrat der AP-Stelle und entfernt den Zucker-Phosphat-Rest. Die DNA-Polymerase I stellt den beschädigten Strang wieder her.

Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.