Das Nukleosom Es ist die Grundeinheit der DNA-Verpackung in eukaryotischen Organismen. Es ist daher das kleinste Chromatinkompressionselement.
Das Nukleosom ist als Oktamer von Proteinen aufgebaut, die als Histone bezeichnet werden, oder als trommelförmige Struktur, auf die etwa 140 nt DNA gewickelt sind, was fast zwei vollständige Umdrehungen ergibt..
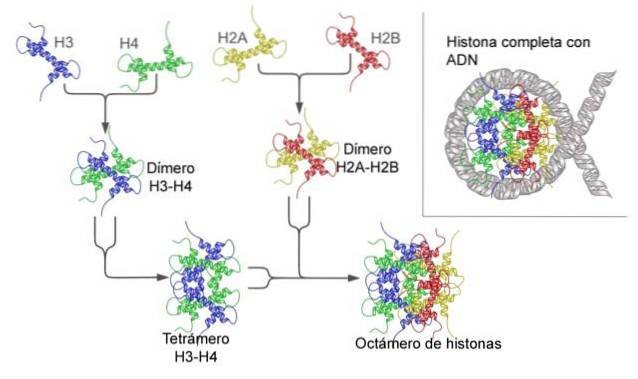
Zusätzlich werden zusätzliche 40-80 nt DNA als Teil des Nukleosoms angesehen, und es ist die DNA-Fraktion, die eine physikalische Kontinuität zwischen einem Nukleosom und einem anderen in komplexeren Chromatinstrukturen (wie der 30-nm-Chromatinfaser) ermöglicht..
Der Histoncode war eines der ersten molekular verstandenen epigenetischen Kontrollelemente..
Artikelverzeichnis
Nukleosomen ermöglichen:
Im einfachsten Sinne bestehen Nukleosomen aus DNA und Proteinen. DNA kann praktisch jede Doppelkern-DNA sein, die im Kern der eukaryotischen Zelle vorhanden ist, während alle nukleosomalen Proteine zu dem Satz von Proteinen gehören, die Histone genannt werden..
Histone sind kleine Proteine mit einer hohen Beladung an basischen Aminosäureresten; Dies ermöglicht es, der hohen negativen Ladung der DNA entgegenzuwirken und eine effiziente physikalische Wechselwirkung zwischen den beiden Molekülen herzustellen, ohne die Steifheit der kovalenten chemischen Bindung zu erreichen..
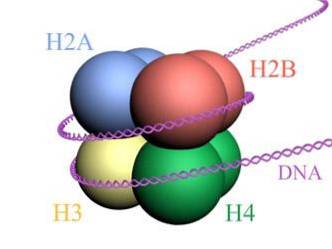
Histone bilden ein trommelartiges Oktamer mit zwei Kopien oder Monomeren der Histone H2A, H2B, H3 und H4. Die DNA macht fast zwei vollständige Umdrehungen an den Seiten des Oktamers und fährt dann mit einem Teil der Linker-DNA fort, die mit Histon H1 assoziiert ist, um zurückzukehren, um zwei vollständige Umdrehungen an einem anderen Histonoctamer zu ergeben..
Der Octamer-Satz, die assoziierte DNA und die entsprechende Linker-DNA ist ein Nukleosom.
Genomische DNA besteht aus extrem langen Molekülen (mehr als ein Meter beim Menschen unter Berücksichtigung aller Chromosomen), die in einem extrem kleinen Kern verdichtet und organisiert werden müssen.
Der erste Schritt bei dieser Verdichtung erfolgt durch Bildung von Nukleosomen. Allein mit diesem Schritt wird die DNA etwa 75-mal verdichtet.
Dies führt zu einer linearen Faser, aus der nachfolgende Niveaus der Chromatinverdichtung aufgebaut werden: die 30-nm-Faser, die Schleifen und die Schleifen von Schleifen..
Wenn sich eine Zelle entweder durch Mitose oder durch Meiose teilt, ist der endgültige Verdichtungsgrad das mitotische bzw. meiotische Chromosom selbst..
Die Tatsache, dass Histonoctamere und DNA elektrostatisch interagieren, erklärt teilweise ihre effektive Assoziation, ohne die Fließfähigkeit zu verlieren, die erforderlich ist, um Nukleosomen zu dynamischen Elementen für die Verdichtung und Zersetzung von Chromatin zu machen..
Es gibt jedoch ein noch überraschenderes Wechselwirkungselement: Die N-terminalen Enden der Histone liegen außerhalb des Inneren des Oktamers frei, das kompakter und inerter ist..
Diese Enden interagieren nicht nur physikalisch mit der DNA, sondern unterliegen auch einer Reihe von kovalenten Modifikationen, von denen der Verdichtungsgrad des Chromatins und die Expression der assoziierten DNA abhängen..
Die Menge der kovalenten Modifikationen, unter anderem in Bezug auf Typ und Anzahl, wird zusammen als Histoncode bezeichnet. Diese Modifikationen umfassen Phosphorylierung, Methylierung, Acetylierung, Ubiquitinierung und Sumoylierung von Arginin- und Lysinresten an den N-Termini von Histonen..
Jede Veränderung bestimmt zusammen mit anderen innerhalb desselben Moleküls oder in Resten anderer Histone, insbesondere der Histone H3, die Expression oder Nicht-Expression der assoziierten DNA sowie den Verdichtungsgrad des Chromatins..
In der Regel wurde beispielsweise festgestellt, dass hypermethylierte und hypoacetylierte Histone bestimmen, dass die assoziierte DNA nicht exprimiert wird und dass Chromatin in einem kompakteren Zustand vorliegt (heterochromatisch und daher inaktiv)..
Im Gegensatz dazu ist euchromatische DNA (weniger kompakt und genetisch aktiv) mit einem Chromatin assoziiert, dessen Histone hyperacetyliert und hypomethyliert sind..
Wir haben bereits gesehen, dass der kovalente Modifikationsstatus von Histonen den Expressionsgrad und die lokale Chromatinverdichtung bestimmen kann. Auf globaler Ebene wird die Chromatinverdichtung auch durch kovalente Modifikationen von Histonen in Nukleosomen reguliert.
Es wurde zum Beispiel gezeigt, dass konstitutives Heterochromatin (das niemals exprimiert wird und dicht gepackt ist) dazu neigt, an der Kernschicht zu haften, wodurch die Kernporen frei bleiben..
Konstitutives Euchromatin (das immer exprimiert wird, wie dasjenige, das Gene zur Erhaltung der Zellen enthält und sich in Regionen mit laxem Chromatin befindet) tut dies in großen Schleifen, die die zu transkribierende DNA der Transkriptionsmaschinerie aussetzen.
Andere Regionen genomischer DNA oszillieren zwischen diesen beiden Zuständen in Abhängigkeit von der Entwicklungszeit des Organismus, den Wachstumsbedingungen, der Zellidentität usw..
Um ihren Plan für die Entwicklung, Expression und Erhaltung von Zellen zu erfüllen, müssen die Genome eukaryotischer Organismen genau regulieren, wann und wie sich ihre genetischen Möglichkeiten manifestieren müssen..
Ausgehend von den in ihren Genen gespeicherten Informationen befinden sich diese im Kern in bestimmten Regionen, die ihren Transkriptionszustand bestimmen..
Wir können daher sagen, dass eine andere der grundlegenden Rollen von Nukleosomen durch die Veränderungen im Chromatin, die es zu definieren hilft, die Organisation oder Architektur des Kerns ist, in dem sie sich befinden..
Diese Architektur ist vererbt und dank der Existenz dieser modularen Elemente der Informationsverpackung phylogenetisch erhalten.



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.