Das Mikrofilamente oder Aktinfilamente sind eine der drei Grundkomponenten des Zytoskeletts eukaryotischer Zellen (Mikrofilamente, Mikrotubuli und Zwischenfilamente) und bestehen aus kleinen Filamenten eines Proteins namens Actin (Aktinpolymere)..
Bei Eukaryoten sind die Gene, die für Aktin-Mikrofilamente kodieren, in allen Organismen hoch konserviert, weshalb sie häufig als molekulare Marker für Studien verschiedener Art verwendet werden.

Mikrofilamente sind im gesamten Cytosol verteilt, kommen jedoch besonders häufig in der Region vor, die unter der Plasmamembran liegt, wo sie ein komplexes Netzwerk bilden und sich mit anderen speziellen Proteinen verbinden, um das Cytoskelett zu bilden..
Mikrofilamentnetzwerke im Zytoplasma von Säugetierzellen werden von zwei der sechs für Actin beschriebenen Gene kodiert, die an der Dynamik der Mikrofilamente beteiligt sind und sogar bei der Stammzelldifferenzierung von großer Bedeutung sind.
Viele Autoren sind sich einig, dass Mikrofilamente die vielfältigsten, vielseitigsten und wichtigsten Proteine im Zytoskelett der meisten eukaryotischen Zellen sind, und es ist wichtig zu bedenken, dass diese nicht in prokaryotischen Mikroorganismen vorkommen..
In diesem Zelltyp gibt es andererseits Filamente, die zu Mikrofilamenten homolog sind, aber aus einem anderen Protein bestehen: dem MreB-Protein..
Gegenwärtig wird angenommen, dass das für dieses Protein kodierende Gen ein mögliches Ahnengen für eukaryotisches Aktin ist. Die Sequenzhomologie der Aminosäuren, aus denen das MreB-Protein besteht, beträgt jedoch nur 15% in Bezug auf die Aktinsequenz.
Da sie ein wesentlicher Bestandteil des Zytoskeletts sind, kann jeder phänotypische Defekt sowohl in Mikrotubuli als auch in Zwischenfilamenten und Aktin-Mikrofilamenten (Zytoskelett) unterschiedliche zelluläre und systemische Pathologien verursachen.
Artikelverzeichnis
Mikrofilamente bestehen aus Proteinmonomeren der Aktinfamilie, die in eukaryotischen Zellen sehr häufig kontraktile Proteine enthalten, da sie auch an der Muskelkontraktion beteiligt sind..
Diese Filamente haben einen Durchmesser zwischen 5 und 7 nm, daher werden sie auch als dünne Filamente bezeichnet und bestehen aus zwei Formen von Aktin: der Kugelform (G-Aktin) und der Filamentform (F-Aktin)..
Die Proteine, die am Zytoskelett beteiligt sind, sind als γ- und β-Actine bekannt, während diejenigen, die an der Kontraktion beteiligt sind, normalerweise α-Actine sind..
Der Anteil von globulärem Actin und filamentösem Actin im Cytosol hängt von den zellulären Bedürfnissen ab, da Mikrofilamente sehr variable und vielseitige Strukturen sind, die durch Polymerisation und Depolymerisation ständig wachsen und sich verkürzen..
G-Actin ist ein kleines globuläres Protein, das aus fast 400 Aminosäuren besteht und ein Molekulargewicht von etwa 43 kDa aufweist.
Die G-Actin-Monomere, aus denen die Mikrofilamente bestehen, sind in Form eines helikalen Strangs angeordnet, da jedes eine Verdrehung erfährt, wenn es mit dem nächsten assoziiert wird.
G-Actin assoziiert mit einem Ca2 + -Molekül und einem ATP-Molekül, die seine Kugelform stabilisieren. während F-Actin nach Hydrolyse des terminalen Phosphats des ATP-Moleküls zu G-Actin erhalten wird, was zur Polymerisation beiträgt.
Aktinfilamente können in "Bündeln" oder "Netzwerken" organisiert werden, die innerhalb von Zellen unterschiedliche Funktionen haben. Die Bündel bilden parallele Strukturen, die durch ziemlich starre Querbrücken verbunden sind.
Netzwerke hingegen sind lockerere Strukturen wie dreidimensionale Netze mit den Eigenschaften von halbfesten Gelen..
Es gibt viele Proteine, die mit Aktinfilamenten oder Mikrofilamenten assoziiert sind und als ABP (aus dem Englischen) bekannt sind Aktin-bindende Proteine), die bestimmte Websites dafür haben.
Viele dieser Proteine ermöglichen es Mikrofilamenten, mit den beiden anderen Komponenten des Zytoskeletts zu interagieren: Mikrotubuli und Zwischenfilamente sowie mit den anderen Komponenten der Innenseite der Plasmamembran..
Andere Proteine, mit denen Mikrofilamente interagieren, umfassen Kernschichten und Spektrin (in roten Blutkörperchen).
Da globuläre Aktinmonomere immer auf die gleiche Weise und in die gleiche Richtung ausgerichtet binden, haben Mikrofilamente eine definierte Polarität mit zwei Enden: einem "mehr" und einem "weniger"..
Die Polarität dieser Filamente ist sehr wichtig, da sie an ihrem positiven Ende, wo die neuen G-Actin-Monomere hinzugefügt werden, erheblich schneller wachsen..

Das erste, was während der Polymerisation von Aktinfilamenten stattfindet, ist ein Prozess, der als "Keimbildung" bekannt ist und aus der Assoziation von drei Monomeren des Proteins besteht.
Diesem Trimer werden an beiden Enden neue Monomere zugesetzt, so dass das Filament wächst. G-Actin-Monomere sind in der Lage, ATP bei jeder Bindung zu hydrolysieren, was Auswirkungen auf die Polymerisationsgeschwindigkeit hat, da Actin-ATP-Einheiten schwieriger dissoziieren als Actin-ADP-Einheiten..
ATP ist für die Polymerisation nicht erforderlich und die spezifische Rolle seiner Hydrolyse wurde noch nicht aufgeklärt..
Einige Autoren sind der Ansicht, dass das mit diesen Prozessen verbundene ATP bis zu 40% des gesamten Zellumsatzes dieses Energiemoleküls ausmachen kann, da Aktinpolymerisationsereignisse schnell reversibel sind.
Sowohl die Polymerisation von Aktinfilamenten als auch deren Depolymerisation sind Prozesse, die durch eine Reihe spezifischer Proteine stark reguliert werden, die für die Umgestaltung der Filamente verantwortlich sind..
Ein Beispiel für die Proteine, die die Depolymerisation regulieren, ist der Aktin-Depolymerisationsfaktor Cofilin. Ein anderes Protein, Profilin, hat eine entgegengesetzte Funktion, da es die Assoziation von Monomeren stimuliert (indem es den Austausch von ADP gegen ATP stimuliert)..
Mikrofilamente interagieren mit Myosinfilamenten, die mit Transmembranproteinen assoziiert sind, die eine Domäne im Cytosol und eine andere im Zelläußeren haben, und sind so an den Prozessen der Zellmobilität beteiligt.
Diese mit der Plasmamembran assoziierten Mikrofilamente vermitteln verschiedene zelluläre Reaktionen auf verschiedene Klassen von Stimuli. Beispielsweise wird die Zelladhäsion in Epithelgeweben durch Transmembranproteine gesteuert, die als Cadherine bekannt sind und mit Mikrofilamenten interagieren, um Antwortfaktoren zu rekrutieren..
Aktinfilamente interagieren mit Zwischenfilamenten, um zu bewirken, dass extrazelluläre Stimuli an Schlüsselstellen wie Ribosomen und Chromosomen innerhalb des Kerns übertragen werden..
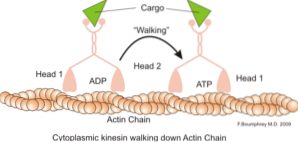
Eine klassische und viel untersuchte Funktion von Mikrofilamenten ist ihre Fähigkeit, "Brücken", "Schienen" oder "Autobahnen" für die Bewegung des Motorproteins Myosin I zu bilden, das Transportvesikel von Organellen auf das Membranplasma im Sekret laden kann Wege.
Mikrofilamente interagieren auch mit Myosin II, um den kontraktilen Ring zu bilden, der sich während der Zytokinese bildet, genau während des letzten Stadiums der Zellteilung, in dem das Zytosol von Stamm- und Tochterzellen getrennt wird..
Im Allgemeinen modulieren F-Actin-Mikrofilamente die Verteilung einiger Organellen wie des Golgi-Komplexes, des endoplasmatischen Retikulums und der Mitochondrien. Darüber hinaus sind sie auch an der räumlichen Positionierung von mRNAs beteiligt, so dass sie von Ribosomen gelesen werden..
Der gesamte zelluläre Satz von Mikrofilamenten, insbesondere diejenigen, die eng mit der Plasmamembran verwandt sind, ist an der Bildung der wellenförmigen Membranen von Zellen beteiligt, die eine konstante aktive Bewegung aufweisen.
Sie sind auch an der Bildung von Mikrovilli und anderen häufigen Beulen auf der Oberfläche vieler Zellen beteiligt..
Mikrofilamente sind am Gallensekretionsprozess in Hepatozyten (Leberzellen) sowie an peristaltischen Bewegungen (koordinierte Kontraktion) der Leberkanäle beteiligt.
Sie tragen aufgrund ihrer Assoziation mit verschiedenen cytosolischen Elementen und der Kontrolle, die sie über die Topographie dieser intrazellulären Elemente ausüben, zur Differenzierung von Plasmamembrandomänen bei..
Es gibt nur wenige Krankheiten, die mit primären Defekten in der Struktur oder mit regulatorischen Proteinen und Enzymen bei der Synthese von Mikrofilamenten verbunden sind, obwohl diese direkt an einer Vielzahl von Funktionen beteiligt sind..
Die geringe Rate an Krankheiten und Missbildungen in der Primärstruktur von Mikrofilamenten beruht auf der Tatsache, dass es im Allgemeinen mehrere Gene gibt, die sowohl für Actin als auch für seine regulatorischen Proteine kodieren, ein Phänomen, das als "genetische Redundanz" bekannt ist..
Eine der am besten untersuchten Pathologien ist die Verglasung der Eizellen auf ihrem Zytoskelett, bei der eine Unterbrechung des Netzwerks kortikaler Mikrofilamente sowie eine Depolymerisation und Desorganisation der Mikrotubuli der mitotischen Spindel beobachtet wird..
Im Allgemeinen verursacht diese Verglasung eine chromosomale Dispersion, da sie zu einer Störung der Verdichtung des gesamten Chromatins führt.
Die Zellen, die eine größere Organisation und einen größeren Anteil an Mikrofilamenten in ihrem Zytoskelett haben, sind die Zellen des gestreiften Muskels, daher sind die meisten Pathologien mit einer Fehlfunktion des kontraktilen Apparats verbunden.
Defekte oder atypische Mikrofilamente wurden auch mit der als Paget-Krankheit bekannten Knochenerkrankung in Verbindung gebracht..


Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.