Calmodulin ist ein Begriff, der "Calcium-moduliertes Protein" bedeutet und sich auf ein kleines intrazelluläres Protein bezieht, das die Eigenschaft hat, an das Calciumion (Ca ++) zu binden und viele seiner intrazellulären Wirkungen zu vermitteln. Der Ursprung des Wortes liegt in der Kombination der englischen Wörter "Calcium", "moduliert" und "Protein", die sich zusammen ergeben LIMETTEcium MODULated proteIM.
Unter den Mineralelementen, die Teil der Konstitution tierischer Organismen werden, ist Kalzium, gefolgt von Phosphor, bei weitem am häufigsten, da Knochen durch Ablagerung großer Mengen von Mineralsalzen, die aus diesem Ion gebildet werden, in seiner Matrix gebildet wird.

Natürlich sind diese Mineralsalze von Kalzium für die Konstitution und Konformation des Skelettsystems von Wirbeltieren essentiell, aber es ist die ionisierte Form von Kalzium (Ca ++) in Lösung in Körperflüssigkeiten, die eine relevante physiologische Bedeutung für das Leben von Kalzium erhält Tiere. Organismen.
Dieses Kation mit zwei überschüssigen positiven elektrischen Ladungen in seiner Struktur kann als Stromträger wirken, indem es sich durch die Zellmembran bewegt und sein elektrisches Potential in vielen erregbaren Körperzellen, hauptsächlich im Herzmuskel, verändert.
Von größerer physiologischer Relevanz ist jedoch die Tatsache, dass viele zelluläre Regulationsreaktionen, die durch externe Stimuli wie Neurotransmitter, Hormone oder andere physikalische oder biochemische Faktoren ausgelöst werden, Arten von Stoffwechselkaskaden sind, an denen mehrere Proteine nacheinander beteiligt sind, von denen einige Enzyme erfordern Kalzium zur Aktivierung oder Inaktivierung.
In diesen Fällen wird dann gesagt, dass Kalzium als zweiter Botenstoff in einer Stoffwechselkaskade fungiert, die dazu bestimmt ist, ein Endergebnis zu erzielen, das der zellulären Reaktion entspricht, die erforderlich ist, um ein auf einem anderen Niveau als dem der Zelle selbst erkanntes Bedürfnis zu befriedigen. und dass es von ihr diese besondere Antwort erfordert.
Calcium kann direkt auf sein biochemisches Ziel einwirken, um seine Aktivität zu beeinflussen, erfordert jedoch häufig die Beteiligung eines Proteins, an das es binden muss, um seine Wirkung auf das zu modifizierende Protein (die zu modifizierenden Proteine) auszuüben. Calmodulin ist eines dieser Mediatorproteine.
Artikelverzeichnis
Calmodulin, hoch allgegenwärtig, da es in fast allen Zelltypen eukaryotischer Organismen exprimiert wird, ist ein kleines saures Protein mit einem Molekulargewicht von etwa 17 kDa, dessen Struktur unter den Arten hoch konserviert ist..
Es ist ein monomeres Protein, das heißt, es besteht aus einer einzelnen Polypeptidkette, die an ihren terminalen Enden die Form von globulären Domänen hat, die durch eine Alpha-Helix verbunden sind. Jede globuläre Domäne hat zwei Motive, die als EF-Hand bekannt sind. EF Hand), die typisch für calciumbindende Proteine sind.
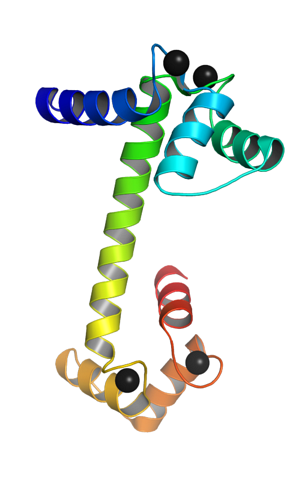
Diese topologischen Motive der "EF-Hand" repräsentieren eine Art übergeordnete Strukturen; sind in jeder globulären Domäne durch eine Region von großer Flexibilität miteinander verbunden, und in jeder von ihnen gibt es eine Bindungsstelle für Ca ++, die insgesamt 4 Stellen für jedes Calmodulinmolekül ergibt.
Die Bindung positiv geladener Calciumionen wird durch das Vorhandensein von Aminosäureresten mit negativ geladenen Seitenketten an den Calciumbindungsstellen von Calmodulin ermöglicht. Diese Rückstände sind drei Aspartate und ein Glutamat..
Alle bisher für Calmodulin bekannten Funktionen sind in einer Gruppe von Wirkungen zusammengefasst, die durch den Anstieg des zytosolischen Kalziums gefördert werden, das durch seinen Eintritt aus dem extrazellulären Raum oder seinen Austritt aus den intrazellulären Ablagerungen erzeugt wird: Mitochondrien und endoplasmatisches Retikulum.
Viele der Wirkungen von Kalzium werden von diesem Ion ausgeführt, indem es direkt auf seine Zielproteine einwirkt, die verschiedene Arten und Funktionen haben können. Einige dieser Proteine können nicht direkt beeinflusst werden, benötigen jedoch Calcium, um an Calmodulin zu binden, und es ist dieser Komplex, der auf das vom Ion beeinflusste Protein einwirkt..
Diese Zielproteine sollen von Calcium-Calmodulin abhängig sein und Dutzende von Enzymen umfassen, wie Proteinkinasen, Proteinphosphatasen, Nukleotidcyclasen und Phosphodiesterasen; alle an einer Vielzahl von physiologischen Funktionen beteiligt, einschließlich:
- Stoffwechsel
- Partikeltransport
- Viszerale Mobilität
- Die Sekretion von Substanzen
- Befruchtung der Eizellen
- Genexpression
- Zellvermehrung
- Die strukturelle Integrität von Zellen
- Interzelluläre Kommunikation usw..
Zu den Calmodulin-abhängigen Proteinkinasen gehören: Myosin-Leichtkettenkinase (MLCK), Phosphorylasekinase und Ca ++ / Calmodulinkinasen I, II und III.
Somit wird die durch Kalziumsignale codierte Information (Zunahme oder Abnahme ihrer intrazellulären Konzentration) durch dieses und andere Kalzium-bindende Proteine "decodiert", die die Signale in biochemische Veränderungen umwandeln; Mit anderen Worten, Calmodulin ist ein Zwischenprotein in calciumabhängigen Signalprozessen.
Calmodulin ist ein sehr vielseitiges Protein, da seine "Ziel" -Proteine in Form, Sequenz, Größe und Funktion sehr unterschiedlich sind. Da es sich um ein Protein handelt, das als „Sensor“ für Calciumionen fungiert, hängt sein Wirkungsmechanismus von den Änderungen ab, die in seiner Struktur und / oder Konformation induziert werden, sobald es an vier dieser Ionen bindet..
Die Wirkmechanismen lassen sich anhand einer kurzen Überprüfung der Beteiligung an einigen physiologischen Prozessen wie der Kontraktion der glatten viszeralen Muskulatur und der Anpassung an Gerüche der Haarzellen der Riechschleimhaut in der Nase veranschaulichen..
Die Kontraktion der Skelett- und Herzmuskulatur wird ausgelöst, wenn der Anstieg des zytosolischen Ca ++ Werte über 10-6 mol / l erreicht und dieses Ion an Troponin C bindet, das allosterische Veränderungen erfährt, die Tropomyosin beeinflussen. Tropomyosin bewegt sich wiederum und legt seine Myosin-Bindungsstellen in Actin frei, was dazu führt, dass der kontraktile Prozess ausgelöst wird.
Troponin C existiert nicht in glatten Muskeln und der Anstieg von Ca ++ über das angegebene Niveau fördert seine Bindung an Calmodulin. Der Ca-Calmodulinkomplex aktiviert die Kinase der leichten Kette von Myosin (MLCK), die wiederum diese leichte Kette phosphoryliert, Myosin aktiviert und den kontraktilen Prozess auslöst..
Der Anstieg von Ca ++ erfolgt durch seinen Eintritt von außen oder seinen Austritt aus dem sarkoplasmatischen Retikulum durch die Wirkung von Inositoltriphosphat (IP3), das von Phospholipase C in der Kaskade freigesetzt wird, die durch an Gq-Protein gekoppelte Rezeptoren aktiviert wird. Eine Entspannung tritt auf, wenn Ca ++ durch die Wirkung von Transportern aus dem Cytosol entfernt wird und zu seinen Ursprungsorten zurückkehrt.
Ein wichtiger Unterschied zwischen beiden Arten der Kontraktion besteht darin, dass Ca ++ in gestreiften Muskeln (Herz und Skelett) allosterische Veränderungen durch Bindung an sein Protein Troponin induziert, während in glatten Muskeln die durch Ca-Calmodulin hervorgerufenen Veränderungen kovalent sind und die Phosphorylierung von Myosin beinhalten.
Sobald die Wirkung von Ca ++ beendet ist, ist daher die Teilnahme eines anderen Enzyms erforderlich, um das von der Kinase hinzugefügte Phosphat zu entfernen. Dieses neue Enzym ist die Myosin-Leichtkettenphosphatase (MLCP), deren Aktivität nicht von Calmodulin abhängt, sondern über andere Wege reguliert wird.
In der Realität hört der kontraktile Prozess der glatten Muskulatur nicht vollständig auf, aber der Grad der Kontraktion bleibt aufgrund des Gleichgewichts der Wirkungen beider Enzyme, der durch Ca ++ und Calmodulin kontrollierten MLCK und der MLCP auf einem mittleren Niveau anderen behördlichen Kontrollen unterworfen.
Das Geruchsempfinden wird ausgelöst, wenn Geruchsrezeptoren in den Zilien von Zellen auf der Oberfläche der Geruchsschleimhaut aktiviert werden..
Diese Rezeptoren sind an ein heterotrimeres G-Protein gekoppelt, das als "Golf" (olfaktorisches G-Protein) bekannt ist und drei Untereinheiten aufweist: "αolf", "ß" und "γ"..
Wenn olfaktorische Rezeptoren als Reaktion auf einen Geruch aktiviert werden, dissoziieren die Untereinheiten dieses Proteins und die "αolf" -Untereinheit aktiviert das Enzym Adenylcyclase und produziert cyclisches Adenosinmonophosphat (cAMP)..
CAMP aktiviert CNG-ähnliche Kanäle (aktiviert durch cyclische Nukleotide) für Calcium und Natrium. Diese Ionen treten in die Zelle ein, depolarisieren sie und lösen Aktionspotentiale aus, deren Frequenz die Intensität des Geruchs bestimmt..
Das eintretende Kalzium, das dazu neigt, die Zelle zu depolarisieren, hat einen antagonistischen Effekt der negativen Rückkopplung, etwas später, indem es an Calmodulin bindet und zwischen ihnen den Kanal schließt und den depolarisierenden Reiz eliminiert, obwohl der Geruchsstimulus anhält. Dies wird als Sensoranpassung bezeichnet.
Pflanzen reagieren auch auf Unterschiede in der intrazellulären Konzentration von Calciumionen durch das Protein Calmodulin. In diesen Organismen teilen Calmoduline viele strukturelle und funktionelle Eigenschaften mit ihren Gegenstücken bei Tieren und Hefen, obwohl sie sich in einigen funktionellen Aspekten unterscheiden..
Beispielsweise bindet Calmodulin in Pflanzen an kurze Peptidsequenzen innerhalb ihrer Zielproteine und induziert strukturelle Veränderungen, die ihre Aktivitäten als Reaktion auf interne Variationen von Calcium verändern..
Inwieweit Calmodulin Prozesse kontrolliert, die denen bei Tieren in Pflanzen entsprechen, ist bis heute umstritten..


Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.