Das Glykolipide Sie sind Membranlipide mit Kohlenhydraten in ihren polaren Kopfgruppen. Sie weisen die asymmetrischste Verteilung unter den Membranlipiden auf, da sie ausschließlich in der äußeren Monoschicht von Zellmembranen vorkommen und in der Plasmamembran besonders häufig vorkommen.
Wie die meisten Membranlipide haben Glykolipide eine hydrophobe Region, die aus unpolaren Kohlenwasserstoffschwänzen besteht, und eine Kopf- oder Polarregion, die je nach fraglichem Glykolipid aus verschiedenen Molekülklassen bestehen kann..
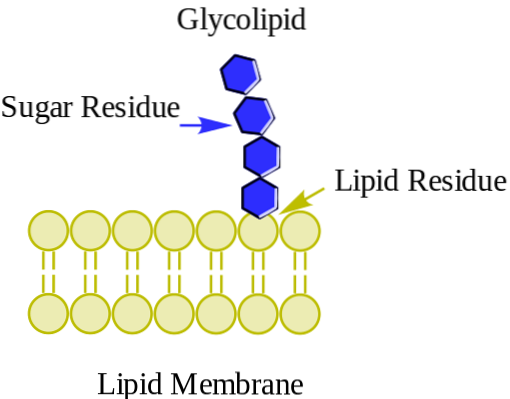
Glykolipide kommen in einzelligen Organismen wie Bakterien und Hefen sowie in Organismen vor, die so komplex sind wie Tiere und Pflanzen.
In tierischen Zellen bestehen Glykolipide überwiegend aus einem Sphingosingerüst, während in Pflanzen die beiden häufigsten Diglyceride und Sulfonsäurederivate entsprechen. In Bakterien gibt es auch Glycosylglyceride und Derivate von acylierten Zuckern.
In Pflanzen sind Glykolipide in den Chloroplastenmembranen konzentriert, während sie in Tieren in der Plasmamembran reichlich vorhanden sind. Glykolipide sind zusammen mit Glykoproteinen und Proteoglykanen ein wichtiger Bestandteil des Glykokalyx, der für viele zelluläre Prozesse von entscheidender Bedeutung ist.
Glykolipide, insbesondere solche tierischer Zellen, neigen dazu, sich durch Wasserstoffbrücken zwischen ihren Kohlenhydrateinheiten und durch Van-der-Waals-Kräfte zwischen ihren Fettsäureketten miteinander zu verbinden. Diese Lipide sind in Membranstrukturen vorhanden, die als Lipidflöße bekannt sind und mehrere Funktionen haben.
Die Funktionen von Glykolipiden sind vielfältig, aber bei Eukaryoten ist ihre Position auf der Außenseite der Plasmamembran unter verschiedenen Gesichtspunkten relevant, insbesondere bei Kommunikations-, Adhäsions- und Zelldifferenzierungsprozessen..
Artikelverzeichnis
Glykolipide sind Glykokonjugate, die eine sehr heterogene Gruppe von Molekülen bilden, deren gemeinsames Merkmal das Vorhandensein von Saccharidresten ist, die durch glucosidische Bindungen an eine hydrophobe Einheit gebunden sind, die Acylglycerin, Ceramid oder Prenylphosphat sein kann.
Seine Klassifizierung basiert auf dem Molekülgerüst, das die Verbindung zwischen der hydrophoben und der polaren Region darstellt. Abhängig von der Identität dieser Gruppe haben wir also:
Diese Glykolipide haben wie Glycerolipide ein Diacylglycerin- oder Monoalkylmonoacylglycerin-Grundgerüst, an das die Zuckerreste durch glucosidische Bindungen gebunden sind..
Glycoglycerolipide sind hinsichtlich ihrer Kohlenhydratzusammensetzung relativ einheitlich, und Galactose- oder Glucosereste können in ihrer Struktur gefunden werden, von der ihre Hauptklassifikation abgeleitet ist, nämlich:
Diese Lipide haben als "Skelett" -Molekül einen Teil Ceramid, an den verschiedene Fettsäuremoleküle gebunden sein können..
Sie sind hochvariable Lipide, nicht nur hinsichtlich der Zusammensetzung ihrer hydrophoben Ketten, sondern auch hinsichtlich der Kohlenhydratreste in ihrem Polarkopf. Sie sind in zahlreichen Säugetiergeweben reichlich vorhanden.
Ihre Klassifizierung basiert eher auf der Art der Substitution oder der Identität des Saccharidanteils als auf der Region, die aus den hydrophoben Ketten besteht. Entsprechend den Substitutionsarten ist die Klassifizierung dieser Sphingolipide wie folgt:
Neutrale Glucosphingolipide: diejenigen, die im Saccharidteil Hexosen, N-Acetylhexosamine und Methylpentosen enthalten.
Sulfatide: Sie sind die Glucosphingolipide, die Sulfatester enthalten. Sie sind negativ geladen und kommen besonders häufig in den Myelinscheiden von Gehirnzellen vor. Die häufigsten haben einen Galaktoserest.
Ganglioside: Auch als Sialosylglykolipide bekannt, sind sie solche, die Sialinsäure enthalten, weshalb sie auch als saure Glycosphingolipide bekannt sind.
Phosphoinositido-Glykolipide: Das Skelett besteht aus Phosphoinositido-Ceramiden.
Sie sind Lipide, die normalerweise als stabile Anker für Proteine in der Lipiddoppelschicht erkannt werden. Sie werden posttranslational an das C-terminale Ende vieler Proteine angefügt, die typischerweise der Außenseite der cytoplasmatischen Membran zugewandt sind..
Sie bestehen aus einem Glucanzentrum, einem Phospholipidschwanz und einem Phosphoethanolaminanteil, der sie verbindet..
Glykolipide können Saccharideinheiten aufweisen, die durch N- oder O-Glucosidbindungen und sogar durch nichtglucosidische Bindungen wie Ester- oder Amidbindungen an das Molekül gebunden sind..
Der Saccharidanteil ist nicht nur in der Struktur, sondern auch in der Zusammensetzung sehr variabel. Dieser Saccharidanteil kann aus Mono-, Di-, Oligo- oder Polysacchariden verschiedener Typen bestehen. Sie können Aminozucker und sogar saure, einfache oder verzweigte Zucker enthalten.
Hier ist eine kurze Beschreibung der allgemeinen Struktur der drei Hauptklassen von Glykolipiden:
Wie zuvor erwähnt, können Glycoglycerolipide bei Tieren Galactose- oder Glucosereste aufweisen, ob phosphatiert oder nicht. Die Fettsäureketten in diesen Lipiden liegen zwischen 16 und 20 Kohlenstoffatomen.
In Galactoglycerolipiden erfolgt die Vereinigung zwischen dem Zucker und dem Lipidrückgrat durch β-glucosidische Bindungen zwischen dem C-1 von Galactose und dem C-3 von Glycerin. Die anderen beiden Kohlenstoffe von Glycerin werden entweder mit Fettsäuren verestert oder C1 wird durch eine Alkylgruppe und C2 durch eine Acylgruppe substituiert..
Normalerweise wird ein einzelner Galactoserest beobachtet, obwohl über die Existenz von Digalactoglycerolipiden berichtet wurde. Wenn es sich um ein Slufogalactoglycerolipid handelt, befindet sich die Sulfatgruppe normalerweise an C-3 des Galactoserests.
Die Struktur von Glycerolipiden ist etwas anders, insbesondere in Bezug auf die Anzahl der Glucosereste, bei denen es sich um bis zu 8 Reste handeln kann, die durch Bindungen vom α (1-6) -Typ miteinander verbunden sind. Das Glucosemolekül, das das Lipidgerüst überbrückt, ist über eine α (1-3) -Bindung an dieses gebunden..
In Sulfoglycoglycerolipiden ist die Sulfatgruppe an Position 6 des terminalen Glucoserests an den Kohlenstoff gebunden.
Wie die anderen Sphingolipide stammen Glycosphingolipide von einem L-Serin, das mit einer langkettigen Fettsäure kondensiert ist und eine Sphingoidbase bildet, die als Sphingosin bekannt ist. Wenn eine andere Fettsäure an Kohlenstoff 2 von Sphingosin bindet, entsteht ein Ceramid, das die gemeinsame Basis für alle Sphingolipide ist.
Je nach Art des Sphingolipids bestehen diese aus D-Glucose-, D-Galactose-, N-Acetyl-D-Galactosamin- und N-Acetylglucosamin-Resten sowie Sialinsäure. Ganglioside sind hinsichtlich der Auswirkungen der Oligosaccharidketten möglicherweise die vielfältigsten und komplexesten.
In diesen Glykolipiden können die Reste des Glucanzentrums (Glucosamin und Mannose) durch Zugabe von Phosphoethanolamingruppen und anderen Zuckern auf unterschiedliche Weise modifiziert werden. Diese Sorte bietet ihnen eine große strukturelle Komplexität, die für ihre Einführung in die Membran wichtig ist..
Die Chloroplasten vieler Algen und höherer Pflanzen sind mit neutralen Galactoglycerolipiden angereichert, deren Eigenschaften denen von Cerebrosiden bei Tieren ähneln. Mono- und Digalactolipide sind an eine Diglycerideinheit β-gebunden, während Sulfolipide nur von α-Glucose abgeleitet sind..
In Bakterien sind Glycosylglyceride strukturell analog zu tierischen Phosphoglyceriden, enthalten jedoch Kohlenhydratreste, die durch Glycosylierung an der 3-Position von sn-1,2-Diglycerid verbunden sind. Acylierte Zuckerderivate enthalten kein Glycerin, sondern direkt an Zucker gebundene Fettsäuren.
Die häufigsten Saccharidreste unter bakteriellen Glykolipiden sind Galactose, Glucose und Mannose..
Bei Tieren spielen Glykolipide unter anderem eine wichtige Rolle bei der Zellkommunikation, Differenzierung und Proliferation, Onkogenese, elektrischen Abstoßung (bei polaren Glykolipiden) und Zelladhäsion..
Seine Anwesenheit in vielen Zellmembranen von Tieren, Pflanzen und Mikroorganismen erklärt seine wichtige Funktion, die insbesondere mit den Eigenschaften multifunktionaler Lipidflöße zusammenhängt..
Der Kohlenhydratanteil der Glycosphingolipide ist eine Determinante für die Antigenität und Immunogenität der Zellen, die sie tragen. Es kann an interzellulären Erkennungsprozessen sowie an zellulären "sozialen" Aktivitäten beteiligt sein..
Galactoglycerolipide in Pflanzen spielen aufgrund ihrer relativen Häufigkeit in Pflanzenmembranen eine wichtige Rolle bei der Festlegung von Membraneigenschaften wie Stabilität und funktioneller Aktivität vieler Membranproteine..
Die Rolle von Glykolipiden in Bakterien ist ebenfalls vielfältig. Einige der Glycoglycerolipide sind erforderlich, um die Stabilität der Doppelschicht zu verbessern. Sie dienen auch als Vorläufer für andere Membrankomponenten und unterstützen auch das Wachstum von Anoxie oder Phosphatmangel..
GPI-Anker oder Glucosidylphosphatidylinositole sind auch in Lipidflößen vorhanden, sind an der Signalübertragung, an der Pathogenese vieler parasitärer Mikroorganismen und an der Orientierung der apikalen Membran beteiligt.
Man kann dann sagen, dass die allgemeinen Funktionen von Glykolipiden sowohl in Pflanzen, Tieren als auch in Bakterien der Herstellung der Stabilität und Fließfähigkeit der Membran entsprechen; Teilnahme an spezifischen Lipid-Protein-Wechselwirkungen und Zellerkennung.
1. Abdel-mawgoud, A. M. & Stephanopoulos, G. (2017). Einfache Glykolipide von Mikroben: Chemie, biologische Aktivität und Stoffwechseltechnik. Synthetische und Systembiotechnologie, 1-17.
2. B. Alberts, A. Johnson, J. Lewis, D. Morgan, M. Raff, K. Roberts & P. Walter (2015). Molekularbiologie der Zelle (6. Aufl.). New York: Garland Science.
3. Ando, T., Imamura, A., Ishida, H. & Kiso, M. (2007). Synthese von Glykolipiden. Carbohydrate Research, 797 & ndash; 813.
4. Benson, A. (1964). Pflanzenmembranlipide. Annu. Rev. Plant. Physiol., 15, 1 & ndash; 16.
5. Bronislaw, L., Liau, Y. U. N. H. & Slomiany, A. (1987). Tierische Glykoglycerolipide. Prog. Lipid Res., 26, 29 & ndash; 51.
6. Holzl, G. & Dormann, P. (2007). Struktur und Funktion von Glycoglycerolipiden in Pflanzen und Bakterien. Prog. Lipid Res., 46, 225 & ndash; 243.
7. Honke, K. (2013). Biosynthese und biologische Funktion von Sulfoglycolipiden. Proc. Jpn. Acad. Ser. B, 89 (4), 129 & ndash; 138.
8. Kanfer, J. & Hakomori, S. (1983). Sphingolipid-Biochemie. (D. Hanahan, Hrsg.), Handbook of Lipid Research 3 (1. Aufl.).
9. Koynova, R. & Caffrey, M. (1994). Phasen und Phasenübergänge der Glycoglycerolipide. Chemistry and Physics of Lipids, 69, 181 & ndash; 207.
10. Law, J. (1960). Glykolipide. Annual Reviews, 29, 131-150.
11. Paulick, M. G. & Bertozzi, C. R. (2008). Der Glycosylphosphatidylinositol-Anker: Eine komplexe Membranverankerung. Biochemistry, 47, 6991 & ndash; 7000.



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.