
Das postmendelsche Vererbung Es enthält alle Ideen und / oder Werke, die nach den 1865 vom religiösen Naturforscher Gregorio Mendel veröffentlichten Studien entstanden sind und die genetische Phänomene oder Verhaltensweisen erklären, die sich von den hier beschriebenen unterscheiden.
Gregorio Mendel gilt als Vater der Genetik, da er vor mehr als 100 Jahren durch die Untersuchung von Kreuzungen zwischen Erbsenpflanzen die ersten Erbgesetze erlassen hat. Das erste Gesetz, "Gesetz der Segregation" genannt, schlägt vor, dass Gene in verschiedenen oder alternativen Formen existieren können.

Diese alternativen Formen werden "Allele" genannt und beeinflussen direkt die phänotypischen Eigenschaften einer Art. Allele können Haarfarbe, Augenfarbe, Haartyp usw. bestimmen..
In seinem ersten Gesetz schlug Mendel vor, dass jedes phänotypische Merkmal, das von den Eltern auf ihre Kinder übertragen wird, von zwei Allelen kontrolliert wird, die jeweils von einem Elternteil stammen (eines von der „Mutter“ und eines vom „Vater“)..
Wenn die Eltern verschiedene Allele beisteuern, ergibt sich demnach eine Mischung, bei der eines der Allele dominiert (das eine, das im Phänotyp ausgedrückt wird) und das andere rezessiv ist (es wird nicht gesehen oder ist "latent" ")..
Mendels zweites Gesetz ist als "Gesetz der unabhängigen Vererbung" bekannt, da Mendel in seinen Kreuzen feststellte, dass das Vererbungsmuster, das die Allele charakterisierte, die ein Merkmal definierten, das Vererbungsmuster der Allele, die ein anderes Merkmal definierten, nicht beeinflusste.
Artikelverzeichnis
Seit Gregor Mendel vor mehr als 100 Jahren seine berühmten Gesetze vorschlug, wurden neue Mechanismen aufgeklärt, um die Vererbung von Charakteren zu erklären, von denen viele als widersprüchlich oder erheblich von den von Mendel beschriebenen angesehen werden können..
Ein Beispiel für die von der "modernen" Wissenschaft beschriebenen Phänomene, deren Fortschritte nur dank der Weiterentwicklung experimenteller Strategien und Forschungsmethoden möglich waren, ist unter anderem die Vererbung in Verbindung mit Geschlecht, unvollständiger Dominanz und Kodominanz..
Geschlechtsgebundene Vererbung hat mit Genen zu tun, die nur auf den Geschlechtschromosomen gefunden werden. Beim Menschen wird das Geschlecht durch die X- und Y-Chromosomen bestimmt. Frauen haben zwei Kopien von X (XX) und Männer haben eine von X und eine von Y (XY)..
Die Gene, die auf jedem dieser beiden Chromosomen gefunden werden, tragen zu den Merkmalen bei, die mit dem Geschlecht verbunden sind. Gene auf dem X-Chromosom sind bei Männern und Frauen vorhanden, Y-Gene jedoch nur bei Männern.
Die geschlechtsgebundene Vererbung wurde zum ersten Mal von William Bateson beobachtet und beschrieben, einem der ersten Wissenschaftler, der Mendels Arbeit „wiederentdeckte“ und sie nutzte, um auf der Grundlage seiner Schlussfolgerungen neue Experimente durchzuführen..
Bateson fertigte experimentelle Hühnerkreuze mit Gefieder in verschiedenen Farben an, einige mit dunklen und glatten Federn, andere mit glatten Federn und mit einem Streifenmuster..
Durch diese Kreuze stellte er fest, dass die resultierenden Männchen bei den Nachkommen immer ein Gefieder mit einem Streifenmuster hatten.
Erst in der dritten Generation, als ein gestreiftes Männchen mit einer Henne mit dunklen und glatten Federn gekreuzt wurde, wurden einige Hähne mit schwarzem Gefieder erhalten. Dieser resultierende Anteil der Kreuze widersprach den von Mendel vorgeschlagenen Anteilen.
Nach Batesons Arbeit fand Thomas Henry Morgan eine logische Erklärung für die seltsamen Ergebnisse von Bateson, indem er den Unterschied in der Färbung bei Hühnern auf die Tatsache zurückführte, dass das Streifenmuster von einem dominanten Allel codiert wurde, das auf dem X-Chromosom gefunden wurde..
Bei Hühnern entspricht im Gegensatz zum Menschen das heterogametische Geschlecht (XY) den Weibchen, während das homogametische Geschlecht den Männchen entspricht (XX)..
Das von Bateson und Morgan untersuchte Allel wurde dann von allen Männern mit gestreiftem Gefieder exprimiert, aber wenn ein Mann zwei X-Chromosomen mit dem der schwarzen Färbung entsprechenden Allel erbte, erschienen Männer mit dunklem Gefieder und ohne Streifenmuster..
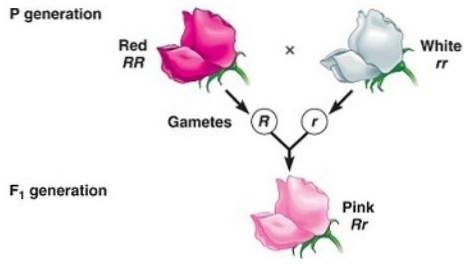
Eine unvollständige Dominanz liegt vor, wenn der bei den Nachkommen beobachtete Phänotyp, der sich aus der Kreuzung zweier homozygoter Individuen (eines dominant und das andere rezessiv) ergibt, ein "intermediärer" Phänotyp zwischen dem dominanten und dem rezessiven ist..
Es hat mit der "Unfähigkeit" des "dominanten" Allels zu tun, die Expression des rezessiven Allels vollständig zu maskieren, weshalb der resultierende Phänotyp einer Mischung der von beiden Allelen codierten Merkmale entspricht, die oft als "verdünnt" bezeichnet werden Phänotyp “der Dominante.
Zum Beispiel in den Dondiego-Pflanzen bei Nacht (Mirabilis Jalapa) Die typische Farbe von Wildblumenblättern ist rot. Wenn eine Pflanze mit roten Blütenblättern mit einer reinen Linienpflanze mit weißen Blütenblättern gekreuzt wird, wird erhalten, dass alle Nachkommen rosa (rosa) Blütenblätter sind.
Wenn zwei dominante homozygote Individuen gekreuzt werden und bei den Nachkommen ein dritter Phänotyp beobachtet wird, der den beiden durch die beiden elterlichen Allele bestimmten Merkmalen entspricht, spricht man von Codominanz.
In diesem Fall würde der "dritte Phänotyp", wenn man mit einem Beispiel fortfährt, das dem der Blüten im vorherigen Beispiel ähnlich ist, nicht rosa Blüten entsprechen, sondern einer Pflanze mit weißen Blüten und roten Blüten auf demselben Stamm, da die beiden Allele "codominate", und keiner maskiert den anderen.
Oft sind Codominanz und unvollständige Dominanz jedoch phänotypisch schwer zu unterscheiden, da beide einen Phänotyp eines Individuums erzeugen, der zwei verschiedene Allele zu exprimieren scheint..
Genotypisch unterscheiden sich jedoch die Individuen, die codominante Allele präsentieren, von denen, die eine unvollständige Dominanz aufweisen, da in einem Fall die beiden dominanten Allele in der gleichen Menge exprimiert werden und in dem anderen Fall die Anteile ungleich sind..
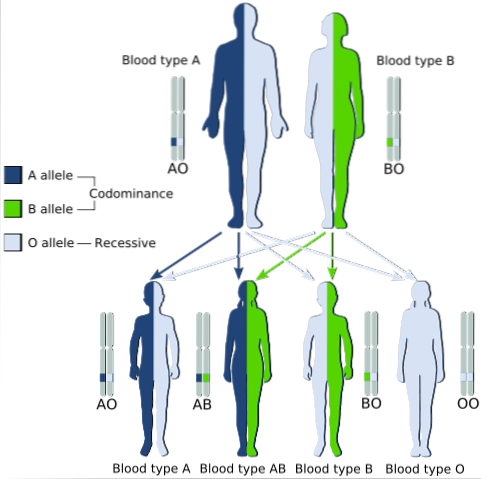
Ein klassisches Beispiel für Codominanz ist die Art der Blutgruppe beim Menschen, da die Allele, die für die Antigene A und B in menschlichen Populationen kodieren, codominant sind.
Das heißt, wenn die Allele A und B zusammen in den Zellen eines Individuums gefunden werden, hat er die Blutgruppe AB.
Sie sind rezessive Allele für einen Charakter, der, wenn er in homozygoter Form gefunden wird, für das Individuum tödlich ist. Personen, die diese Allele präsentieren, werden nicht geboren, da sie das normale Funktionieren der molekularen Maschinerie und die Translation von Genen verhindern.
Viele Stellen im Genom eines Individuums interagieren mit anderen Teilen desselben Genoms, dh die spezifische Modifikation eines Allels innerhalb des Genoms kann die Expression anderer Allele innerhalb der Zellen eines Individuums direkt konditionieren.
Diese Wechselwirkungen sind schwer zu bestimmen und werden in zwei verschiedene Typen eingeteilt: Gene, die auf demselben genetischen Weg interagieren, und solche, die auf verschiedenen genetischen Wegen interagieren..
Ein klassisches und bekanntes Beispiel für Gene, die auf verschiedenen genetischen Wegen interagieren, hat mit den Genen zu tun, die die Hautfarbe von Kornnattern bestimmen (Pantherophis guttatus), deren gemeinsame Farbe ein sich wiederholendes Muster aus schwarzen und orangefarbenen Flecken ist.

Dieser Phänotyp wird von zwei verschiedenen Pigmenten erzeugt, die einer strengen genetischen Kontrolle unterliegen. Ein Gen bestimmt die orange Farbe und hat zwei Allele: ein O +, das für das Pigment kodiert, und ein anderes O-, das die vollständige Abwesenheit des orangefarbenen Pigments bestimmt..
Ein anderes anderes Gen bestimmt das Vorhandensein oder Fehlen des schwarzen Pigments. Die Allele dieses Gens sind als B + bekannt, dasjenige, das das Vorhandensein des schwarzen Pigments bestimmt, und B-, das das Fehlen des Pigments bestimmt.
Beide Gene sind nicht miteinander verbunden und bestimmen unabhängig voneinander die Farbe von Schlangen, dh eine Schlange mit dem O + / B + -Genotyp ist eine normale Schlange mit einem orange-schwarzen Phänotyp, aber jede genotypische Variation führt zu einem anderen Phänotyp..
Das klassische Beispiel für diese Gene ist dasjenige, das für die „Glockenblumen“ -Blumen (aus der Familie der Campanulaceae) beschrieben wurde, deren „wilder“ Phänotyp Pflanzen mit blauen Blüten entspricht..
Die blaue Farbe von Bluebell-Blüten wird nicht durch ein Gen bestimmt, sondern durch einen Stoffwechselweg, bei dem mehrere Enzyme, die jeweils von einem anderen Gen kodiert werden, an der Synthese eines Pigments beteiligt sind..
Somit gibt es Phänotypen, die dadurch gekennzeichnet sind, dass sie das Ergebnis der Interaktion mehrerer Gene sind, deren Produkte am gleichen Stoffwechselweg beteiligt sind und bei denen die Mutation oder Fehlfunktion eines dieser Gene einen Phänotyp von weißen, depigmentierten Blüten ergibt, in diesem Fall..



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.