Das oxidative Phosphorylierung ist ein Prozess, bei dem ATP-Moleküle aus ADP und P synthetisiert werdenich (anorganisches Phosphat). Dieser Mechanismus wird von Bakterien und eukaryotischen Zellen ausgeführt. In eukaryotischen Zellen findet die Phosphorylierung in der mitochondrialen Matrix nicht-photosynthetischer Zellen statt.
Die ATP-Produktion wird durch den Elektronentransfer von den Coenzymen NADH oder FADH angetriebenzwei zum O.zwei. Dieser Prozess stellt die höchste Energieproduktion in der Zelle dar und wird aus dem Abbau von Kohlenhydraten und Fetten abgeleitet..
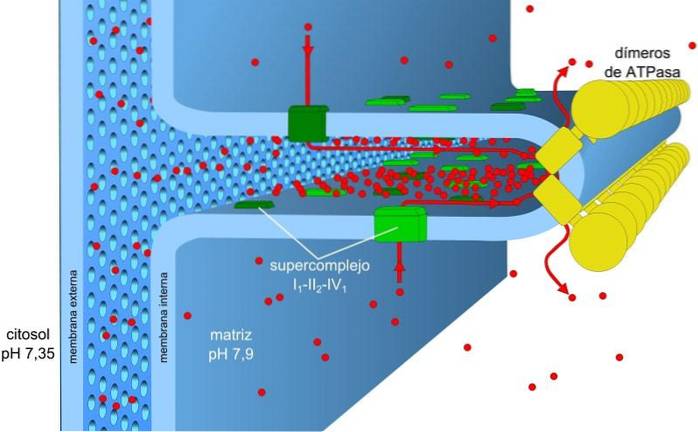
Die in den Ladungs- und pH-Gradienten gespeicherte Energie, auch als Protonenantriebskraft bekannt, ermöglicht diesen Prozess. Der erzeugte Protonengradient bewirkt, dass der äußere Teil der Membran aufgrund der Protonenkonzentration (H) eine positive Ladung aufweist+) und die mitochondriale Matrix ist negativ.
Artikelverzeichnis
Die Prozesse des Elektronentransports und der oxidativen Phosphorylierung sind mit einer Membran verbunden. Bei Prokaryoten werden diese Mechanismen durch die Plasmamembran ausgeführt. In eukaryotischen Zellen assoziieren sie mit der Mitochondrienmembran.
Die Anzahl der in Zellen gefundenen Mitochondrien variiert je nach Zelltyp. Beispielsweise fehlen Erythrozyten bei Säugetieren diese Organellen, während andere Zelltypen wie Muskelzellen bis zu Millionen davon haben können..
Die Mitochondrienmembran besteht aus einer einfachen Außenmembran, einer etwas komplexeren Innenmembran und dazwischen dem Intermembranraum, in dem sich viele ATP-abhängige Enzyme befinden.
Die äußere Membran enthält ein Protein namens Porin, das die Kanäle für die einfache Diffusion kleiner Moleküle bildet. Diese Membran ist für die Aufrechterhaltung der Struktur und Form der Mitochondrien verantwortlich.
Die innere Membran hat eine höhere Dichte und ist reich an Proteinen. Es ist auch für Moleküle und Ionen undurchlässig. Um sie zu durchqueren, benötigen sie Intermembranproteine, um sie zu transportieren..
Innerhalb der Matrix erstrecken sich Falten der inneren Membran und bilden Rippen, die es ihr ermöglichen, eine große Fläche in einem kleinen Volumen zu haben.
Das Mitochondrium gilt als zentraler Produzent zellulärer Energie. Es enthält die Enzyme, die an den Prozessen des Zitronensäurezyklus, der Fettsäureoxidation und der Redoxenzyme sowie der Proteine für den Elektronentransport und die ADP-Phosphorylierung beteiligt sind.
Der Protonenkonzentrationsgradient (pH-Gradient) und der Ladungsgradient oder das elektrische Potential in der inneren Membran der Mitochondrien sind für die Protonenmotivkraft verantwortlich. Die geringe Permeabilität der inneren Membran für Ionen (außer H.+) ermöglicht den Mitochondrien einen stabilen Spannungsgradienten.
Elektronischer Transport, Protonenpumpen und ATP-Gewinnung erfolgen dank der Protonenmotivkraft gleichzeitig in den Mitochondrien. Der pH-Gradient hält saure Bedingungen in der Intermembran und der mitochondrialen Matrix unter alkalischen Bedingungen aufrecht.
Für jeweils zwei auf O übertragene Elektronenzwei Etwa 10 Protonen werden durch die Membran gepumpt, wodurch ein elektrochemischer Gradient entsteht. Die dabei freigesetzte Energie wird allmählich durch den Durchgang von Elektronen durch die Transportkette erzeugt.
Die Energie, die während der Oxidations-Reduktions-Reaktionen von NADH und FADH freigesetzt wirdzwei ist beträchtlich hoch (etwa 53 kcal / mol für jedes Elektronenpaar). Um bei der Herstellung von ATP-Molekülen verwendet zu werden, muss es schrittweise mit dem Durchgang von Elektronen durch Transporter erzeugt werden.
Diese sind in vier Komplexe organisiert, die sich auf der inneren Mitochondrienmembran befinden. Die Kopplung dieser Reaktionen an die ATP-Synthese erfolgt in einem fünften Komplex.
NADH überträgt ein Elektronenpaar, das in den Komplex I der Elektronentransportkette eintritt. Elektronen werden über einen Eisen-Schwefel-Transporter auf das Flavin-Mononukleotid und dann auf Ubichinon (Coenzym Q) übertragen. Dieser Prozess setzt eine große Energiemenge frei (16,6 kcal / mol)..
Ubichinon transportiert Elektronen über die Membran zum Komplex III. In diesem Komplex passieren die Elektronen die Cytochrome b und c1 dank eines Eisen-Schwefel-Transporters.
Elektronen wandern von Komplex III zu Komplex IV (Cytochrom-C-Oxidase) und werden einzeln in Cytochrom-C (peripheres Membranprotein) übertragen. In Komplex IV passieren die Elektronen ein Paar Kupferionen (Cuzuzwei+), gefolgt von Cytochrom czu, dann zu einem anderen Paar Kupferionen (Cubzwei+) und von diesem zu Cytochrom a3.
Schließlich werden die Elektronen auf O übertragenzwei Das ist der letzte Akzeptor und bildet ein Wassermolekül (H.zweiO) für jedes empfangene Elektronenpaar. Der Durchgang von Elektronen vom Komplex IV nach O.zwei Es erzeugt auch eine große Menge an freier Energie (25,8 kcal / mol)..
Der Komplex II (Succinat-CoQ-Reduktase) erhält durch Oxidation eines Succinatmoleküls zu Fumarat ein Elektronenpaar aus dem Zitronensäurezyklus. Diese Elektronen werden auf das FAD übertragen und gelangen dann über eine Eisen-Schwefel-Gruppe zu Ubichinon. Von diesem Coenzym gelangen sie zum Komplex III und folgen dem zuvor beschriebenen Weg.
Die Energie, die bei der Elektronentransferreaktion auf das FAD freigesetzt wird, reicht nicht aus, um die Protonen durch die Membran zu treiben, so dass in diesem Schritt der Kette keine Protonenantriebskraft erzeugt wird und folglich das FADH weniger H ergibt+ dass der NADH.
Die Energie, die bei dem zuvor beschriebenen Elektronentransportprozess erzeugt wird, muss zur Herstellung von ATP verwendet werden können, einer Reaktion, die durch das Enzym ATP-Synthase oder -Komplex V katalysiert wird. Die Erhaltung dieser Energie ist als Energiekopplung bekannt, und der Mechanismus wurde schwer zu charakterisieren.
Es wurden mehrere Hypothesen beschrieben, um diese Energieübertragung zu beschreiben. Am besten akzeptiert wird die nachstehend beschriebene chemiosmotische Kopplungshypothese..
Dieser Mechanismus legt nahe, dass die für die ATP-Synthese verwendete Energie von einem Protonengradienten in Zellmembranen stammt. Dieser Prozess greift in die Mitochondrien, Chloroplasten und Bakterien ein und ist mit dem Transport von Elektronen verbunden.
Die Komplexe I und IV des Elektronentransports wirken als Protonenpumpen. Diese unterliegen Konformationsänderungen, die es ihnen ermöglichen, Protonen in den Intermembranraum zu pumpen. In Komplex IV werden für jedes Elektronenpaar zwei Protonen aus der Membran gepumpt und zwei weitere verbleiben in der Matrix und bilden H.zweiODER.
Ubichinon in Komplex III nimmt Protonen aus den Komplexen I und II auf und setzt sie an die Außenseite der Membran frei. Die Komplexe I und III ermöglichen jeweils den Durchgang von vier Protonen für jedes Paar transportierter Elektronen.
Die mitochondriale Matrix hat eine geringe Protonenkonzentration und ein negatives elektrisches Potential, während der Intermembranraum die inversen Bedingungen darstellt. Der Protonenfluss durch diese Membran stellt den elektrochemischen Gradienten dar, der die für die ATP-Synthese erforderliche Energie (± 5 kcal / mol pro Proton) speichert..
Das Enzym ATP-Synthetase ist der fünfte Komplex, der an der oxidativen Phosphorylierung beteiligt ist. Es ist dafür verantwortlich, die Energie des elektrochemischen Gradienten zur Bildung von ATP zu nutzen.
Dieses Transmembranprotein besteht aus zwei Komponenten: F.0 und F.1. Komponente F.0 ermöglicht die Rückkehr von Protonen in die mitochondriale Matrix, die als Kanal fungiert, und das F.1 katalysiert die ATP-Synthese durch ADP und P.ich, unter Verwendung der Energie dieser Rückkehr.
Der ATP-Synthesevorgang erfordert eine strukturelle Änderung von F.1 und die Montage von Komponenten F.0 und F.1. Protonentranslokation durch F.0 verursacht Konformationsänderungen in drei Untereinheiten von F.1, Dadurch kann es als Rotationsmotor wirken und die Bildung von ATP steuern.
Die Untereinheit, die für die Bindung von ADP an P verantwortlich istich es geht von einem schwachen Zustand (L) zu einem aktiven (T) über. Wenn ATP gebildet wird, geht eine zweite Untereinheit in einen offenen Zustand (O) über, der die Freisetzung dieses Moleküls ermöglicht. Nach der Freigabe von ATP wechselt diese Untereinheit vom offenen Zustand in einen inaktiven Zustand (L)..
ADP- und P-Moleküleich an eine Untereinheit binden, die von einem O-Zustand in einen L-Zustand übergegangen ist.
Die Elektronentransportkette und die Phosphorylierung produzieren ATP-Moleküle. Die Oxidation von NADH erzeugt etwa 52,12 kcal / mol (218 kJ / mol) freie Energie.
Die Gesamtreaktion für die Oxidation von NADH ist:
NADH + 1⁄2 O.zwei +H.+ ↔ H.zweiO + NAD+
Übertragung von Elektronen von NADH und FADHzwei tritt durch verschiedene Komplexe auf, wodurch die Änderung der freien Energie ΔG ° in kleinere Energie- "Pakete" aufgeteilt werden kann, die an die ATP-Synthese gekoppelt sind.
Die Oxidation eines Moleküls NADH erzeugt die Synthese von drei Molekülen ATP. Während der Oxidation eines FADH-Molekülszwei koppelt an die Synthese von zwei ATPs.
Diese Coenzyme stammen aus den Glykolyse- und Zitronensäurezyklusprozessen. Für jedes abgebaute Glucosemolekül produzieren sie je nach Standort der Zellen 36 oder 38 ATP-Moleküle. 36 ATP wird im Gehirn und im Skelettmuskel produziert, während 38 ATP im Muskelgewebe produziert wird..
Alle Organismen, einzellig und mehrzellig, benötigen nur minimale Energie in ihren Zellen, um die Prozesse in ihnen auszuführen, und behalten wiederum lebenswichtige Funktionen im gesamten Organismus bei..
Stoffwechselprozesse erfordern Energie, um stattfinden zu können. Der größte Teil der nutzbaren Energie wird aus dem Abbau von Kohlenhydraten und Fetten gewonnen. Diese Energie stammt aus dem oxidativen Phosphorylierungsprozess.
Die ATP-Verwertungsrate in Zellen steuert ihre Synthese und reguliert aufgrund der Kopplung der oxidativen Phosphorylierung mit der Elektronentransportkette im Allgemeinen auch die Geschwindigkeit des Elektronentransports..
Die oxidative Phosphorylierung unterliegt einer strengen Kontrolle, die sicherstellt, dass ATP nicht schneller erzeugt als verbraucht wird. Es gibt bestimmte Schritte im Prozess des Elektronentransports und der gekoppelten Phosphorylierung, die die Geschwindigkeit der Energieerzeugung regulieren..
Die Hauptwege der Energieerzeugung (zelluläres ATP) sind Glykolyse, Zitronensäurezyklus und oxidative Phosphorylierung. Die koordinierte Steuerung dieser drei Prozesse reguliert die ATP-Synthese.
Die Kontrolle der Phosphorylierung durch das Massenwirkungsverhältnis von ATP hängt vom genauen Beitrag der Elektronen in der Transportkette ab. Dies hängt wiederum von [NADH] / [NAD ab+], die durch die Wirkung der Glykolyse und des Zitronensäurezyklus erhöht gehalten wird.
Diese koordinierte Kontrolle erfolgt durch Regulierung der Glykolysekontrollpunkte (PFK durch Citrat gehemmt) und des Zitronensäurezyklus (Pyruvatdehydrogenase, Citrattapease, Isocitratdehydrogenase und α-Ketoglutaratdehydrogenase)..
Komplex IV (Cytochrom-C-Oxidase) ist ein Enzym, das durch eines seiner Substrate reguliert wird, dh seine Aktivität wird durch reduziertes Cytochrom-C (c) gesteuertzwei+), die wiederum im Gleichgewicht mit dem Konzentrationsverhältnis zwischen [NADH] / [NAD+] und das Massenwirkungsverhältnis von [ATP] / [ADP] + [P.ich]].
Je höher das Verhältnis [NADH] / [NAD+] und senken Sie die [ATP] / [ADP] + [P.ich], je höher die Cytochromkonzentration ist [czwei+] und je größer die Aktivität des Komplexes IV. Dies wird zum Beispiel interpretiert, wenn wir Organismen mit verschiedenen Aktivitäten der Ruhe und der hohen Aktivität vergleichen.
Bei einer Person mit hoher körperlicher Aktivität ist der Verbrauch von ATP und damit seine Hydrolyse zu ADP + P.ich wird sehr hoch sein und einen Unterschied im Massenwirkungsverhältnis erzeugen, der eine Zunahme von [czwei+] und damit eine Erhöhung der ATP-Synthese. Bei einer ruhenden Person tritt die umgekehrte Situation auf.
Letztendlich steigt die Geschwindigkeit der oxidativen Phosphorylierung mit der Konzentration von ADP in den Mitochondrien. Diese Konzentration hängt von den ADP-ATP-Translokatoren ab, die für den Transport von Adeninnukleotiden und P verantwortlich sindich vom Cytosol zur mitochondrialen Matrix.
Die oxidative Phosphorylierung wird durch bestimmte chemische Mittel beeinflusst, die es ermöglichen, den Elektronentransport fortzusetzen, ohne dass eine ADP-Phosphorylierung auftritt, wodurch die Energieerzeugung und -erhaltung entkoppelt wird.
Diese Mittel stimulieren den Sauerstoffverbrauch der Mitochondrien in Abwesenheit von ADP und verursachen auch einen Anstieg der ATP-Hydrolyse. Sie wirken, indem sie einen Vermittler eliminieren oder einen Energiezustand der Elektronentransportkette aufbrechen.
2,4-Dinitrophenol, eine schwache Säure, die durch Mitochondrienmembranen gelangt, ist für die Auflösung des Protonengradienten verantwortlich, da sie auf der sauren Seite an sie binden und auf der basischen Seite freisetzen.
Diese Verbindung wurde als "Diätpille" verwendet, da festgestellt wurde, dass sie eine Erhöhung der Atmung, daher eine Erhöhung der Stoffwechselrate und einen damit verbundenen Gewichtsverlust bewirkt. Es wurde jedoch gezeigt, dass seine negative Wirkung sogar zum Tod führen kann.
Die Ableitung des Protonengradienten erzeugt Wärme. Zellen in braunem Fettgewebe verwenden eine hormonell kontrollierte Entkopplung, um Wärme zu erzeugen. Säugetiere und Neugeborene im Winterschlaf, denen Haare fehlen, bestehen aus diesem Gewebe, das als eine Art Wärmedecke dient..
Verbindungen oder Hemmstoffe verhindern beide den Verbrauch von O.zwei (Elektronentransport) wie die damit verbundene oxidative Phosphorylierung. Diese Mittel verhindern die Bildung von ATP, indem sie die beim elektronischen Transport erzeugte Energie nutzen. Daher stoppt die Transportkette, da der Energieverbrauch nicht verfügbar ist..
Das Antibiotikum Oligomycin wirkt in vielen Bakterien als Phosphorylierungsinhibitor und verhindert die Stimulierung der ADP zur ATP-Synthese.
Es gibt auch Ionophormittel, die mit Kationen wie K fettlösliche Komplexe bilden+ und Na+, und sie passieren mit diesen Kationen die Mitochondrienmembran. Die Mitochondrien nutzen dann die beim Elektronentransport erzeugte Energie, um Kationen zu pumpen, anstatt ATP zu synthetisieren..



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.