Das Erythropoetin, Hämopoetin oder EPA ist ein Glykoprotein mit hormonellen Funktionen (Zytokin), das für die Kontrolle der Proliferation, Differenzierung und des Überlebens der Vorläuferzellen von Erythrozyten oder roten Blutkörperchen im Knochenmark, dh der Erythropoese, verantwortlich ist.
Dieses Protein ist einer der verschiedenen Wachstumsfaktoren, die die hämatopoetischen Prozesse steuern, durch die aus einer kleinen Gruppe pluripotenter Stammzellen die im Blut gefundenen Zellen gebildet werden: sowohl Erythrozyten als auch weiße Blutkörperchen und Lymphozyten. Das heißt, Zellen der myeloischen und lymphoiden Linie.
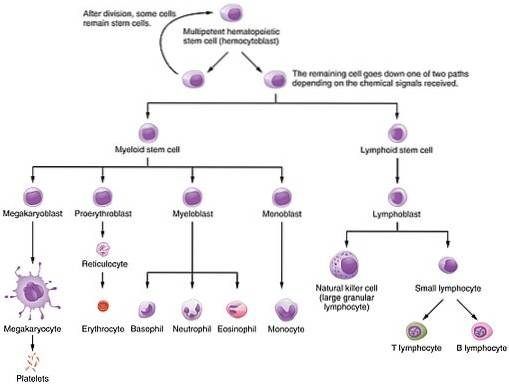
Seine Bedeutung liegt in der funktionellen Bedeutung von Zellen, die zur Vermehrung, Differenzierung und Reifung beitragen, da Erythrozyten für den Sauerstofftransport von der Lunge zu den verschiedenen Geweben des Körpers verantwortlich sind.
Erythropoetin war der erste Wachstumsfaktor, der geklont wurde (1985), und seine Verabreichung zur erfolgreichen Behandlung von Anämie, die durch Nierenversagen verursacht wird, ist derzeit von der American Food and Drug Administration (FDA) zugelassen..
Die Vorstellung, dass die Erythropoese durch einen humoralen Faktor (löslicher Faktor im Kreislauf) gesteuert wird, wurde vor mehr als 100 Jahren von Carnot und Deflandre vorgeschlagen, als sie die positiven Auswirkungen auf den Anstieg des Prozentsatzes der roten Blutkörperchen bei mit Serum behandelten Kaninchen untersuchten. Anämisch Tiere.
Erst 1948 führten Bonsdorff und Jalavisto den Begriff "Erythropoetin" ein, um den humoralen Faktor mit einer spezifischen Auswirkung auf die Produktion von Erythrozyten zu beschreiben..
Artikelverzeichnis
Erythropoetin ist ein Protein aus der Glykoproteinfamilie. Es ist bei sauren pH-Werten stabil und hat ein Molekulargewicht von ca. 34 kDa..
Es hat ungefähr 193 Aminosäuren, einschließlich einer hydrophoben N-terminalen Region mit 27 Resten, die durch co-translationale Verarbeitung entfernt wird; und einen Argininrest an Position 166, der ebenfalls verloren geht, so dass das zirkulierende Protein 165 Aminosäuren aufweist.
In seiner Struktur ist die Bildung von zwei Disulfidbrücken zwischen den in den Positionen 7-161 und 29-33 vorhandenen Cysteinresten zu sehen, die mit seiner Funktionsweise zusammenhängen. Es besteht zu mehr oder weniger als 50% aus Alpha-Helices, die anscheinend an der Bildung einer Kugelregion oder eines kugelförmigen Teils beteiligt sind.
Es enthält 40% Kohlenhydrate, dargestellt durch drei Oligosaccharidketten, die an verschiedene Asparaginsäurereste (Asp) N-gebunden sind, und eine O-Kette, die an einen Serinrest (Ser) gebunden ist. Diese Oligosaccharide bestehen hauptsächlich aus Fucose, Mannose, N-Acetylglucosamin, Galactose und N-Acetylneuraminsäure..
Die Kohlenhydratregion von EPO erfüllt mehrere Rollen:
- Es ist wichtig für seine biologische Aktivität.
- Schützt es vor Abbau oder Beschädigung durch freie Sauerstoffradikale.
- Die Oligosaccharidketten werden für die Sekretion des reifen Proteins benötigt.
Beim Menschen befindet sich das Gen, das für dieses Protein kodiert, in der Mitte des langen Arms von Chromosom 7 in der Region q11-q22; Es befindet sich in einer einzelnen Kopie in einer 5,4-kb-Region und hat fünf Exons und vier Introns. Homologiestudien zeigen, dass seine Sequenz zu 92% mit der anderer Primaten und zu 80% mit der einiger Nagetiere identisch ist..
Während der Entwicklung des Fötus wird Erythropoetin hauptsächlich in der Leber produziert. Es wurde jedoch festgestellt, dass im selben Stadium das Gen, das für dieses Hormon kodiert, auch im mittleren Bereich der Nieren-Nephrone reichlich exprimiert wird..
Nach der Geburt wird das Hormon in allen postnatalen Stadien im Wesentlichen in den Nieren produziert. Insbesondere durch die Zellen der Kortikalis und die Oberfläche der Nierenkörperchen.
Die Leber ist auch an der Produktion von Erythropoetin in den postnatalen Stadien beteiligt, aus denen etwa 20% des gesamten zirkulierenden EPO-Gehalts ausgeschieden werden..
Andere "extra-renale" Organe, in denen eine Erythropoietinproduktion nachgewiesen wurde, umfassen periphere Endothelzellen, glatte Gefäßmuskelzellen und Insulin produzierende Zellen..
Es ist auch bekannt, dass einige EPO-Sekretionszentren im Zentralnervensystem existieren, einschließlich Hippocampus, Cortex, Hirnendothelzellen und Astrozyten..
Die Produktion von Erythropoetin wird nicht direkt durch die Anzahl der roten Blutkörperchen im Blut gesteuert, sondern durch die Zufuhr von Sauerstoff im Gewebe. Ein Sauerstoffmangel im Gewebe stimuliert die Produktion von EPO und seinen Rezeptoren in Leber und Nieren.
Diese Hypoxie-vermittelte Aktivierung der Genexpression ist das Produkt der Aktivierung des Weges einer Familie von Transkriptionsfaktoren, die als Hypoxie-induzierbarer Faktor 1 (HIF-1) bekannt sind. Hypoxie-induzierbarer Faktor 1).
Hypoxie induziert dann die Bildung vieler Proteinkomplexe, die unterschiedliche Funktionen bei der Aktivierung der Erythropoietinexpression erfüllen und direkt oder indirekt an Faktoren binden, die das Aktivierungssignal an den Promotor des EPO-Gens übertragen und dessen Transkription stimulieren..
Andere Stressfaktoren wie Hypoglykämie (niedriger Blutzucker), Anstieg des intrazellulären Kalziums oder das Vorhandensein reaktiver Sauerstoffspezies lösen ebenfalls den HIF-1-Weg aus..
Der Wirkungsmechanismus von Erythropoetin ist recht komplex und hängt hauptsächlich von seiner Fähigkeit ab, verschiedene an der Zellproliferation beteiligte Signalkaskaden zu stimulieren, die wiederum mit der Aktivierung anderer Faktoren und Hormone zusammenhängen.
Im menschlichen Körper eines gesunden Erwachsenen besteht ein Gleichgewicht zwischen der Produktion und Zerstörung von roten Blutkörperchen oder Erythrozyten, und EPO ist an der Aufrechterhaltung dieses Gleichgewichts beteiligt, indem es die verschwindenden Erythrozyten ersetzt.
Wenn die im Gewebe verfügbare Sauerstoffmenge sehr gering ist, nimmt die Expression des für Erythropoetin kodierenden Gens in den Nieren und in der Leber zu. Der Reiz kann auch durch Höhenlagen, Hämolyse, schwere Anämie, Blutungen oder längere Exposition gegenüber Kohlenmonoxid gegeben sein.
Diese Zustände erzeugen einen Hypoxiezustand, der dazu führt, dass die Sekretion von EPO zunimmt, eine größere Anzahl von roten Blutkörperchen produziert wird und der Anteil der im Kreislauf befindlichen Retikulozyten, die eine der Vorläuferzellen von Erythrozyten sind, ebenfalls zunimmt..
Bei der Erythropoese ist EPO hauptsächlich an der Proliferation und Differenzierung von Vorläuferzellen beteiligt, die an der Linie der roten Blutkörperchen beteiligt sind (erythrozytische Vorläufer), aktiviert jedoch auch die Mitose bei Proerythroblasten und basophilen Erythroblasten und beschleunigt auch die Freisetzung von Knochenmarkretikulozyten.
Die erste Ebene, auf der das Protein wirkt, ist die Verhinderung des programmierten Zelltods (Apoptose) der im Knochenmark gebildeten Vorläuferzellen, die durch hemmende Wechselwirkung mit den an diesem Prozess beteiligten Faktoren erreicht wird..
Zellen, die auf Erythropoietin reagieren, besitzen einen spezifischen Rezeptor für Erythropoietin, der als Erythropoietinrezeptor oder EpoR bekannt ist. Sobald das Protein mit seinem Rezeptor einen Komplex bildet, wird das Signal innerhalb der Zelle übertragen: zum Zellkern.
Der erste Schritt für die Signalübertragung ist eine Konformationsänderung, die auftritt, nachdem das Protein an seinen Rezeptor gebunden hat, der gleichzeitig an andere aktivierte Rezeptormoleküle gebunden ist. Unter ihnen ist Janus-Tyrosinkinase 2 (Jack-2).
Unter einigen der Wege, die stromabwärts aktiviert werden, nachdem Jack-2 die Phosphorylierung von Tyrosinresten des EpoR-Rezeptors vermittelt, befindet sich der MAP-Kinase- und Proteinkinase-C-Weg, die Transkriptionsfaktoren aktivieren, die die Expression spezifischer Gene erhöhen.
Wie viele hormonelle Faktoren in Organismen ist Erythropoetin nicht auf eine einzige Funktion beschränkt. Dies wurde durch zahlreiche Untersuchungen aufgeklärt.
Erythropoetin wirkt nicht nur als Proliferations- und Differenzierungsfaktor für Erythrozyten, die für den Transport von Gasen durch den Blutkreislauf wesentlich sind, sondern scheint auch einige zusätzliche Funktionen zu erfüllen, die nicht unbedingt mit der Aktivierung der Zellproliferation und -differenzierung zusammenhängen.
Studien haben gezeigt, dass EPO Zellschäden verhindert, und obwohl seine Wirkmechanismen nicht genau bekannt sind, wird angenommen, dass es apoptotische Prozesse verhindern kann, die durch verringerte oder fehlende Sauerstoffspannung, Erregungstoxizität und Exposition gegenüber freien Radikalen hervorgerufen werden..
Seine Beteiligung an der Prävention von Apoptose wurde durch Wechselwirkung mit bestimmenden Faktoren in den Signalkaskaden untersucht: Janus-Tyrosinkinase 2 (Jak2), Caspase 9, Caspase 1 und Caspase 3, Glykogensynthasekinase-3β, Aktivierungsfaktor von apoptotischen Proteasen 1 (Apaf-1) und andere.
Beteiligt sich an der Hemmung zellulärer Entzündungen durch Hemmung einiger entzündungsfördernder Zytokine wie Interleukin 6 (IL-6), Tumornekrosefaktor Alpha (TNF-α) und Monozyten-Chemo-Lockstoff-Protein 1.
Es wurde gezeigt, dass es im Gefäßsystem bei der Aufrechterhaltung seiner Integrität und bei der Bildung neuer Kapillaren aus vorhandenen Gefäßen in Bereichen ohne Gefäßsystem zusammenarbeitet (Angiogenese). Darüber hinaus verhindert es die Durchlässigkeit der Blut-Hirn-Schranke bei Verletzungen.
Es wird angenommen, dass es die postnatale Neovaskularisation stimuliert, indem die Mobilisierung von Vorläuferzellen vom Knochenmark zum Rest des Körpers erhöht wird..
Es spielt eine wichtige Rolle bei der Entwicklung neuronaler Vorläuferzellen durch die Aktivierung des Kernfaktors KB, der die Produktion von Nervenstammzellen fördert..
In Verbindung mit anderen Zytokinen hat EPO eine "modulierende" Funktion bei der Kontrolle der Proliferations- und Differenzierungswege von Megakaryozyten und Granulozyten-Monozyten..


Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.