Das Erythropoese Es ist der Prozess, durch den rote Blutkörperchen oder Erythrozyten gebildet werden. Diese Blutzellen haben beim Menschen eine durchschnittliche Lebensdauer von 4 Monaten und können sich nicht selbst reproduzieren. Aus diesem Grund müssen neue Erythrozyten geschaffen werden, um diejenigen zu ersetzen, die sterben oder bei Blutungen verloren gehen..
Bei Männern beträgt die Anzahl der roten Blutkörperchen ungefähr 54 Millionen pro Milliliter, während sie bei Frauen etwas geringer ist (48 Millionen). Täglich gehen etwa 10 Millionen Erythrozyten verloren, daher muss eine ähnliche Menge ersetzt werden.
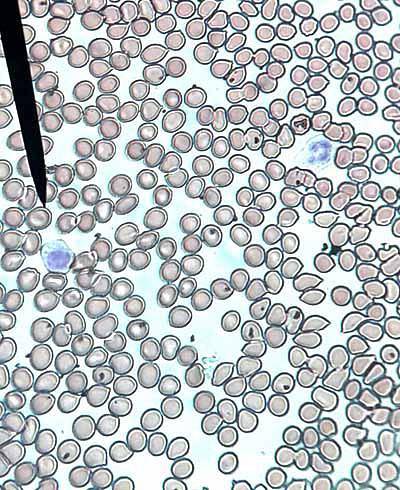
Erythrozyten werden aus kernhaltigen Erythroblasten gebildet, die im roten Knochenmark von Säugetieren vorhanden sind, während sie bei anderen Wirbeltieren hauptsächlich in den Nieren und der Milz produziert werden.
Wenn sie das Ende ihrer Tage erreichen, zersplittern sie; dann umfassen Zellen, die Makrophagen genannt werden, sie. Diese Makrophagen sind in Leber, rotem Knochenmark und Milz vorhanden..
Wenn die roten Blutkörperchen zerstört sind, wird das Eisen recycelt, um wieder verwendet zu werden, während der Rest des Hämoglobins in ein Gallenfarbstoff namens Bilirubin umgewandelt wird..
Die Erythropoese wird durch ein Hormon namens Erythropoietin stimuliert, der Prozess wird jedoch durch verschiedene Faktoren wie Temperatur, Sauerstoffdruck usw. reguliert..
Artikelverzeichnis
Bei erwachsenen Organismen tritt die Erythropoese an spezialisierten Stellen im roten Knochenmark auf, die als erythroblastische Inseln bezeichnet werden. Für die Bildung von Erythrozyten müssen verschiedene Prozesse stattfinden, die von der Zellproliferation bis zur Reifung der roten Blutkörperchen reichen und verschiedene Stadien der Zelldifferenzierung durchlaufen..
Wenn sich Zellen mitotisch teilen, nehmen ihre Größe und die ihres Kerns sowie die Chromatinkondensation und Hämoglobinisierung ab. Außerdem entfernen sie sich vom Herkunftsgebiet.
Im Endstadium verlieren sie den Kern und andere Organellen und gelangen in den Kreislauf, wobei sie durch die zytoplasmatischen Poren von Endothelzellen wandern..
Einige Autoren teilen den gesamten Erythropoeseprozess in zwei Phasen ein, die erste der Zellproliferation und -differenzierung; während andere den Prozess basierend auf spezifischen Eigenschaften der Zelle in jedem Stadium teilen, wenn sie mit Wright-Färbung beobachtet werden. Basierend auf letzterem sind die Stadien der Erythropoese:
Sie sind die ersten Zellen, die gegenüber Erythropoetin empfindlich sind. Einige Autoren nennen sie myeloische Vorläufer oder auch BFU-E, weil sie auf Englisch Akronym sind. Sie zeichnen sich durch die Expression von Oberflächenantigenen wie CD34 sowie durch das Vorhandensein von Erythropoietinrezeptoren in geringen Mengen aus.
Auf Englisch als CFU-E abgekürzt, sind sie in der Lage, kleine Kolonien von Erythroblasten zu produzieren. Ein weiteres Merkmal dieser Zellen ist, dass die Mengen an Erythropoietinrezeptoren viel höher sind als in Burst-Kolonie bildenden Einheiten..
Wird als erstes Reifungsstadium von Erythrozyten angesehen. Sie zeichnen sich durch ihre Größe aus (14 bis 19 µm nach einigen Autoren, bis zu 25 µm nach anderen). Der Kern ist gerundet und enthält auch Nukleolen und reichlich vorhandenes Chromatin.
Wird als erstes Reifungsstadium von Erythrozyten angesehen. Sie zeichnen sich durch ihre Größe aus (14 bis 19 µm nach einigen Autoren, bis zu 25 µm nach anderen). Der Kern ist groß, gerundet, mit Chromatin in Form von Filamenten und 2 oder 3 Nukleolen.
Die Plasma-Eisenaufnahme beginnt in diesem Stadium. Sie haben eine Halbwertszeit von 20 Stunden, um durch Mitose in die nächste Phase zu gelangen.
Sie werden auch Normoblasten genannt und sind kleiner als ihre Vorläufer. Diese Zellen färben sich blau mit lebenswichtiger Färbung, das heißt, sie sind basophil. Der Kern ist kondensiert, die Nukleolen sind verschwunden und sie haben eine große Anzahl von Ribosomen. In diesem Stadium beginnt die Synthese von Hämoglobin.
Zu Beginn sind sie als basophile Erythroblasten vom Typ I bekannt und wandeln sich nach einer mitotischen Teilung in Typ II um, die Basophile bleiben und eine stärkere Hämoglobinsynthese aufweisen. Die ungefähre Dauer beider Zellen zusammen ist ähnlich der von Proerythroblasten.
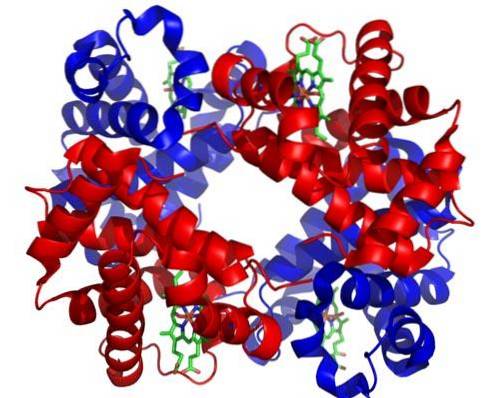
Sie werden durch mitotische Teilung basophiler Erythroblasten vom Typ II gebildet und sind die letzten Zellen, die sich durch Mitose teilen können. Ihre Größe reicht von 8 bis 12 um und sie haben einen abgerundeten und kondensierten Kern.
Das Zytoplasma dieser Zellfärbungen ist mit Wright-Färbung bleigrau. Hat eine hohe Konzentration an Hämoglobin und die Anzahl der Ribosomen bleibt hoch.
6-orthochromatische Erythroblasten
Die Farbe dieser Zellen ist aufgrund der Menge an Hämoglobin, die sie haben, rosa oder rot. Seine Größe ist etwas kleiner als die seiner Vorläufer (7 bis 10 um) und er hat einen kleinen Kern, der durch Exozytose ausgestoßen wird, wenn die Zellen reifen..
Sie entstehen durch Differenzierung orthochromer Erythroblasten, verlieren Organellen und füllen ihr Zytoplasma mit Hämoglobin. Sie bleiben zwei bis drei Tage im roten Knochenmark, bis sie ins Blut wandern, wo sie ihre Reifung abschließen..
Sie sind die reif gebildeten Elemente, das Endprodukt der Erythropoese, die durch die Reifung von Retikulozyten gebildet werden. Sie haben eine bikonkave Form aufgrund des Fehlens eines Kerns und der Wechselwirkung zwischen dem Erythrozyten-Zytoskelett und zwei Proteinen, die als Spectrin und Actin bezeichnet werden.
Sie sind die am häufigsten vorkommenden Blutzellen, sie werden aus Retikulozyten gebildet. Bei Säugetieren haben sie aufgrund des Fehlens eines Kerns und der Wechselwirkung zwischen dem Erythrozyten-Zytoskelett und zwei Proteinen, die als Spectrin und Actin bezeichnet werden, eine bikonkave Form. Bei anderen Wirbeltieren sind sie gerundet und behalten den Kern.
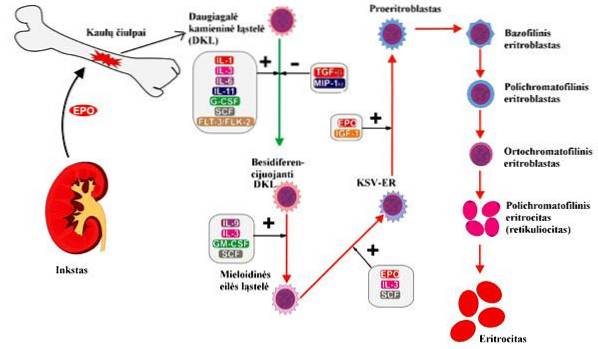
Obwohl Erythropoetin die Bildung roter Blutkörperchen stimuliert, um die Sauerstofftransportkapazität des Blutes zu verbessern, gibt es mehrere grundlegende Mechanismen, um diese Bildung zu regulieren, darunter:
Die Sauerstoffkonzentration im Blut reguliert die Erythropoese. Wenn diese Konzentration im Blutfluss zur Niere sehr niedrig ist, wird die Produktion roter Blutkörperchen stimuliert..
Diese niedrige Konzentration an Gewebe-O2 kann aufgrund von Hypoxämie, Anämie, Nierenischämie oder wenn die Affinität von Hämoglobin für dieses Gas höher als normal ist, auftreten..
Miescher war 1893 der erste, der die Beziehung zwischen Gewebehypoxie und Erythropoese vorschlug. Diese Hypoxie stimuliert jedoch nicht direkt das Knochenmark, um rote Blutkörperchen zu produzieren, wie Miescher vorschlug. Im Gegenteil, es induziert die Niere, das Hormon Erythropoietin zu produzieren.
Die Erythropoietinproduktion aufgrund von Gewebehypoxie ist genetisch reguliert, und die Rezeptoren, die eine solche Hypoxie nachweisen, befinden sich in der Niere. Die Erythropoetin-Produktion ist auch aufgrund eines Abfalls des Sauerstoffpartialdrucks im Gewebe nach Blutungen erhöht..
Die Zellen, die Erythropoetin bilden, befinden sich in Niere und Leber. Die Zunahme der Produktion dieses Hormons während der Anämie ist auf eine Zunahme der Anzahl der Zellen zurückzuführen, die es produzieren..
Testosteron reguliert indirekt die Erythropoese durch Regulierung des Eisenspiegels im Blut. Dieses Hormon wirkt direkt auf die Wirkung eines zytoplasmatischen Proteins namens BMP-Smad (knochenmorphogenetisches Protein-Smad) in Hepatozyten.
Aufgrund der Wirkung von Testosteron wird die Hepcidin-Transkription unterdrückt. Dieses Hepcidin verhindert den Durchgang von Eisen aus Zellen in das Plasma von Makrophagen, die Eisen recyceln, was zu einer drastischen Abnahme des Bluteisens führt.
Wenn eine Hypoferrämie auftritt, kommt es zu einer Hemmung von Erythropoetin, da kein Eisen für die Produktion von Erythrozyten vorhanden ist.
Es wurde gezeigt, dass die Temperatur einen Einfluss auf die Erythripoese hat. Bei sehr niedrigen Temperaturen muss in den Stoffen Wärme erzeugt werden.
Dies erfordert eine Erhöhung der Anzahl der Erythrozyten, um das periphere Gewebe mit Sauerstoff zu versorgen. Es ist jedoch nicht ganz klar, wie diese Art der Regulierung erfolgt..
Anscheinend wird von den Neuronen des Zentralnervensystems Erythropoetin produziert, um sich vor ischämischen Schäden und Apoptose zu schützen. Wissenschaftler konnten dies jedoch noch nicht verifizieren..
Erythropoese-stimulierende Mittel (ESAs) sind Mittel, die für die Stimulierung der Produktion von Erythrozyten verantwortlich sind. Erythropoetin ist das Hormon, das natürlich für diesen Prozess verantwortlich ist, aber es gibt auch synthetische Produkte mit ähnlichen Eigenschaften.
Erythropoetin ist ein Hormon, das hauptsächlich in der Niere synthetisiert wird. In den frühen Entwicklungsstadien ist die Leber auch an der aktiven Produktion von Erythropoetin beteiligt. Mit fortschreitender Entwicklung ist das letztere Gremium jedoch weniger in den Prozess involviert..
Der Erythrozyten beginnt, Rezeptoren für Erythropoetin auf der Membranoberfläche zu dispergieren. Erythropoetin aktiviert eine Reihe von interzellulären Signalübertragungskaskaden, die anfänglich die Hämoglobinsynthese erzeugen und bewirken, dass Retikulozyten schneller wirken und in den Kreislauf freigesetzt werden..
Künstliche ESAs werden je nach Erstellungsdatum und Vermarktungsdatum in Generationen (erste bis dritte) eingeteilt. Strukturell und funktionell ähnlich wie Erythropoetin.
ESAs der ersten Generation sind als Epoetin Alpha, Beta und Delta bekannt. Die ersten beiden werden durch Rekombination aus tierischen Zellen hergestellt und haben im Körper eine Halbwertszeit von ca. 8 Stunden. Das Epoetin-Delta wird unterdessen aus menschlichen Zellen synthetisiert.
Darbepoetin alfa ist eine ESA der zweiten Generation, die aus chinesischen Hamsterzellen unter Verwendung der als rekombinante DNA bezeichneten Technologie hergestellt wird. Es hat eine mehr als dreimal längere Halbwertszeit als ESAs der ersten Generation. Wie bei Epoetinen haben einige Hochleistungssportler Darbepoetin als Mittel zum Doping verwendet.
Continuous Erythropoetin Receptor Activator oder CERA für sein Akronym in Englisch ist der generische Name für ESAs der dritten Generation. Sie versuchen nicht, die Struktur und Funktion von Erythropoetin zu simulieren, sondern stimulieren den Rezeptor für Erythropoetin und verstärken so dessen Wirkung..
Die Halbwertszeit beträgt wie bei den vorherigen Medikamenten mehrere Wochen statt Stunden. Seit 2008 kommerziell genutzt, reicht seine illegale Verwendung bei sportlichen Aktivitäten offenbar zwei oder drei Jahre vor seiner legalen Kommerzialisierung zurück.
Ineffektive oder ineffektive Erythropoese tritt auf, wenn die gebildeten roten Blutkörperchen defekt sind und normalerweise vor oder kurz nach dem Verlassen des Knochenmarks zerstört werden..
Eine ineffektive Erythropoese kann auf Defekte bei der Synthese von Nukleinsäuren, der Hämgruppe oder Globinen zurückzuführen sein. Diese Defekte verursachen verschiedene Arten von Anämie.
In diesem Fall besteht ein Mangel an Folsäure und Cobalamin, die DNA-Synthese wird im Zellkern der Erythrozyten-Promotorzellen gehemmt, so dass sie sich nicht mitotisch teilen können. Das Zytoplasma seinerseits erhöht sein Volumen (Makrozytose), wodurch eine große Zelle namens Megaloblast entsteht.
In diesen Fällen entsteht eine Reihe von Anämien, die als megaloblastische Anämien bezeichnet werden. Die häufigste davon ist die perniziöse Anämie. Bei dieser Krankheit findet keine Aufnahme von Vitamin B12 im Dünndarm statt.
Andere Ursachen für Megaloblastenanämie sind Verdauungskrankheiten, Malabsorption, Folsäuremangel und aufgrund bestimmter Medikamente.
Zu den Symptomen dieser Art von Anämie gehören abnormale Blässe, Reizbarkeit, Appetitlosigkeit, Durchfall, Schwierigkeiten beim Gehen oder Muskelschwäche. Je nach Ursache kann es mit Vitamin- oder Folsäurepräparaten behandelt werden..
Ineffektive Erythropoese aufgrund eines Mangels an Eisensynthese kann zwei Arten von Anämie verursachen; Mikrozytische Anämie mit Eisenmangel und sideroblastische Anämie.
Mikrozytische Anämie ist als eine Gruppe von Anämien bekannt, die durch kleine und hellrote Blutkörperchen gekennzeichnet sind. Sie können unterschiedliche Ursachen haben, einschließlich Thalassämie und ineffektiver Erythropoese.
Eisen- und Hämosiderinspiegel sind bei sideroblastischer Anämie sehr hoch. Hämosiderin ist ein gelbes Pigment, das aus Hämoglobin gewonnen wird und auftritt, wenn der Metallgehalt höher als normal ist. Diese Art von Anämie führt zum Tod von Basophilen im roten Knochenmark und es findet keine Synthese von Hämoglobin statt.
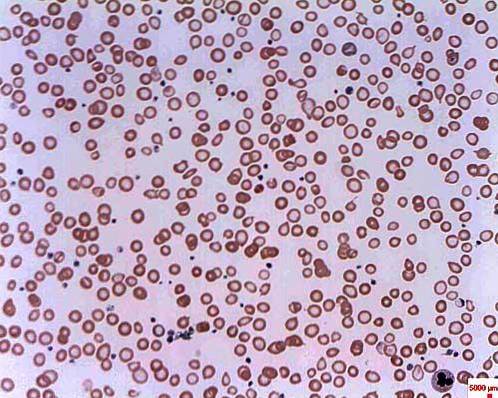
Es wird als sideroblastische Anämie bezeichnet, da sich Erythrozyten aufgrund der Anreicherung von Eisen in Form von Granulaten, die den Namen Sideroblasten erhalten, abnormal entwickeln. Sideroblastische Anämie kann angeboren oder sekundär sein und unterschiedliche Ursachen haben.
In diesem Fall treten Sichelzellenanämie und Beta-Thalassämie auf. Sichelzellenanämie wird auch als Sichelzellenanämie bezeichnet. Es wird durch eine genetische Mutation erzeugt, die zur Substitution von Glutaminsäure durch Valin während der Synthese von Beta-Globin führt.
Aufgrund dieser Substitution nimmt die Affinität von Hämoglobin zu Sauerstoff ab und es kommt zu einer Atrophie des Erythrozyten, die anstelle der normalen bikonkaven Scheibenform eine Sichelform annimmt. Der Patient mit Sichelzellenanämie ist anfällig für Mikroinfarkte und Hämolyse.
Thalassämie ist eine Krankheit, die durch eine unzureichende genetische Kodierung von α- und β-Globinen verursacht wird und zu einem frühen Tod des Erythrozyten führt. Es gibt ungefähr hundert verschiedene Mutationen, die Thalassämie mit unterschiedlichem Schweregrad verursachen können.



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.