Das Zwischenfilamente, in der Literatur auch als "IFs" bekannt (aus dem Englischen Zwischenfilamente) sind eine Familie unlöslicher zytosolischer faseriger Proteine, die in allen Zellen mehrzelliger Eukaryoten vorhanden sind.
Sie sind Teil des Zytoskeletts, eines intrazellulären filamentösen Netzwerks, das hauptsächlich für die Unterstützung der Zellstruktur und für verschiedene metabolische und physiologische Prozesse wie Vesikeltransport, Zellbewegung und -verschiebung usw. verantwortlich ist..
Neben Mikrotubuli und Mikrofilamenten sind Zwischenfilamente an der räumlichen Organisation intrazellulärer Organellen, an den Prozessen der Endozytose und Exozytose sowie an den Prozessen der Zellteilung und der interzellulären Kommunikation beteiligt..
Die ersten Zwischenfilamente, die untersucht und beschrieben wurden, waren Keratine, eine der ersten Arten von Proteinen, deren Struktur in den 1930er Jahren durch Röntgenbeugung analysiert wurde..
Das Konzept der Zwischenfilamente wurde jedoch in den 1980er Jahren von Lazarides eingeführt, der sie als komplexe „mechanische Integratoren des Zellraums“ beschrieb, die sich durch ihre Unlöslichkeit und ihre Fähigkeit zum Zusammenbau auszeichnen. in vitro nach Denaturierung.
Sie werden von vielen Autoren als Stress "Puffer" -Elemente für tierische Zellen angesehen, da sie flexiblere Filamente sind als Mikrotubuli und Mikrofilamente. Sie kommen nicht nur im Zytoskelett vor, sondern sind auch Teil des Nukleoskeletts.
Im Gegensatz zu den anderen faserigen Komponenten des Zytoskeletts sind die Zwischenfilamente nicht direkt an den Prozessen der Zellmobilität beteiligt, sondern wirken sich auf die strukturelle Aufrechterhaltung und den mechanischen Widerstand der Zellen aus..
Artikelverzeichnis
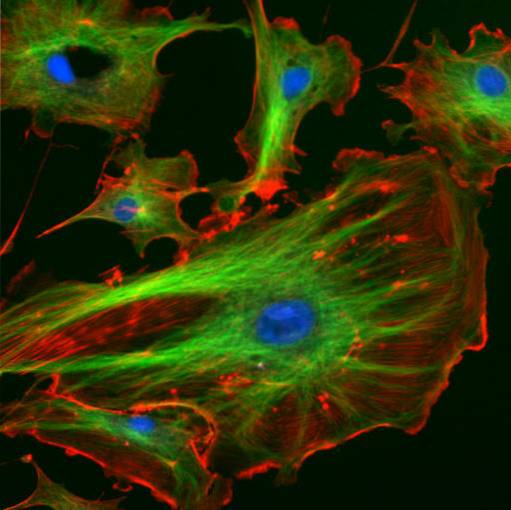
Die Zwischenfilamente haben einen ungefähren Durchmesser von 10 nm, eine strukturelle Eigenschaft, für die sie benannt wurden, da ihre Größe zwischen den Größen liegt, die Myosin- und Actinfilamenten entsprechen, die zwischen 25 bzw. 7 nm liegen.
Sie unterscheiden sich strukturell von den beiden anderen Arten von Zytoskelettfilamenten, bei denen es sich um globuläre Proteinpolymere handelt, darin, dass ihre Bestandteile unterschiedliche α-helikale faserige Proteine mit langer Länge sind, die sich zu seilartigen Strukturen zusammenballen..
Alle Proteine, aus denen die Zwischenfilamente bestehen, haben eine ähnliche molekulare Organisation, die aus einer α-Helix- oder "Seil" -Domäne besteht, die unterschiedliche Mengen an "spulenbildenden" Segmenten gleicher Größe aufweist..
Diese helikale Domäne wird von einem N-terminalen nicht helikalen "Kopf" und einem nicht helikalen "Schwanz" am C-terminalen Ende flankiert, die beide sowohl in der Größe als auch in der Aminosäuresequenz variieren..
Innerhalb der Sequenz dieser beiden Enden sind die Konsensmotive, die für die 6 Arten von Zwischenfilamenten bekannt sind, bekannt.
Bei Wirbeltieren beträgt die "Akkord" -Domäne von cytosolischen Intermediärfilamentproteinen etwa 310 Aminosäurereste, während wirbellose und zytosolische Kernlamina-Proteine etwa 350 Aminosäuren lang sind..
Zwischenfilamente sind „selbstorganisierende“ Strukturen, die keine enzymatische Aktivität besitzen, was sie auch von ihren Gegenstücken im Zytoskelett (Mikrotubuli und Mikrofilamente) unterscheidet..
Diese Strukturen werden zunächst als Tetramere der filamentösen Proteine zusammengesetzt, aus denen sie nur unter dem Einfluss einwertiger Kationen bestehen..
Diese Tetramere sind 62 nm lang und ihre Monomere verbinden sich seitlich miteinander, um "Längeneinheiten" (UFL) zu bilden. Filamente mit Einheitslänge), die als Phase 1 der Baugruppe bekannt ist und sehr schnell erfolgt.
UFLs sind die Vorläufer langer Filamente, und da die Dimere, aus denen sie bestehen, antiparallel und versetzt miteinander verbunden sind, haben diese Einheiten eine zentrale Domäne mit zwei flankierenden Domänen, durch die Phase 2 der Dehnung auftritt, in der die longitudinale Vereinigung von andere UFLs treten auf.
Während der sogenannten Phase 3 der Anordnung tritt eine radiale Verdichtung des Durchmessers der Filamente auf, wodurch reife Zwischenfilamente mit einem Durchmesser von mehr oder weniger 10 nm erzeugt werden..
Die Funktionen der Zwischenfilamente hängen erheblich von der Art der betrachteten Zelle ab, und bei Tieren (einschließlich Menschen) wird ihre Expression gewebespezifisch reguliert, daher hängt sie auch von der Art des untersuchten Gewebes ab.
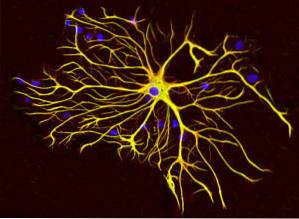
Epithelien, Muskeln, mesenchymale und Gliazellen sowie Neuronen haben verschiedene Arten von Filamenten, die je nach Funktion der Zellen, zu denen sie gehören, spezialisiert sind..
Unter diesen Funktionen sind die strukturelle Aufrechterhaltung der Zellen und die Beständigkeit gegen unterschiedliche mechanische Beanspruchungen am wichtigsten, da diese Strukturen eine gewisse Elastizität aufweisen, die es ihnen ermöglicht, verschiedene Arten von auf die Zellen ausgeübten Kräften abzufedern..
Die Proteine, aus denen die Zwischenfilamente bestehen, gehören zu einer großen und heterogenen Familie filamentöser Proteine, die sich chemisch unterscheiden, aber nach ihrer Sequenzhomologie in sechs Klassen unterteilt sind (I, II, III, IV, V und VI)..
Obwohl es nicht sehr häufig ist, können verschiedene Zelltypen unter ganz bestimmten Bedingungen (Entwicklung, Zelltransformation, Wachstum usw.) mehr als eine Klasse von intermediären filamentbildenden Proteinen coexprimieren.
Keratine machen die meisten Proteine in den Zwischenfilamenten aus, und beim Menschen machen sie mehr als drei Viertel der Zwischenfilamente aus.
Sie haben Molekulargewichte, die zwischen 40 und 70 kDa variieren und sich von anderen Intermediärfilamentproteinen durch ihren hohen Gehalt an Glycin- und Serinresten unterscheiden..
Sie sind aufgrund ihrer isoelektrischen Punkte, die bei sauren Keratinen zwischen 4,9 und 5,4 und bei basischen Keratinen zwischen 6,1 und 7,8 liegen, als saure und basische Keratine bekannt..
In diesen beiden Klassen wurden etwa 30 Proteine beschrieben, die insbesondere in Epithelzellen vorhanden sind, in denen beide Arten von Proteinen „co-polymerisieren“ und zusammengesetzte Filamente bilden..
Viele der Keratine des Zwischenfilaments I finden sich in Strukturen wie Haaren, Nägeln, Hörnern, Federkielen und Krallen, während diejenigen der Klasse II im Cytosol am häufigsten vorkommen.
Desmin ist ein saures Protein mit 53 kDa, das je nach Phosphorylierungsgrad unterschiedliche Varianten aufweist.
Einige Autoren haben Desminfilamente auch als "Zwischenmuskelfilamente" bezeichnet, da ihre Anwesenheit, wenn auch in geringen Mengen, auf alle Arten von Muskelzellen beschränkt ist..
In Myofibrillen befindet sich Desmin in der Z-Linie, weshalb angenommen wird, dass dieses Protein zu den kontraktilen Funktionen der Muskelfasern beiträgt, indem es an der Verbindungsstelle von Myofibrillen und Plasmamembran wirkt..
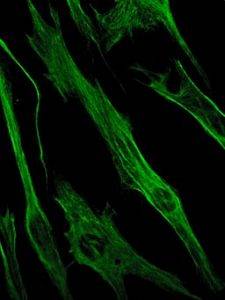
Vimentin ist wiederum ein Protein, das in mesenchymalen Zellen vorhanden ist. Die von diesem Protein gebildeten Zwischenfilamente sind flexibel und widerstehen vielen Konformationsänderungen, die während des Zellzyklus auftreten..
Es kommt in Fibroblasten, glatten Muskelzellen, weißen Blutkörperchen und anderen Zellen des Kreislaufsystems von Tieren vor..
Diese Klasse von Zwischenfilamenten, auch als "Neurofilamente" bekannt, umfasst eines der grundlegenden Strukturelemente neuronaler Axone und Dendriten; werden oft mit den Mikrotubuli assoziiert, aus denen auch diese Strukturen bestehen.
Die Neurofilamente von Wirbeltieren wurden isoliert, wobei festgestellt wurde, dass ein Triplett von Proteinen mit 200, 150 und 68 kDa an der Assemblierung beteiligt ist in vitro.
Sie unterscheiden sich von anderen Zwischenfilamenten dadurch, dass sie seitliche Arme als „Anhänge“ haben, die aus der Peripherie derselben herausragen und in der Wechselwirkung zwischen benachbarten Filamenten und anderen Strukturen funktionieren..
Gliazellen produzieren einen speziellen Typ von Zwischenfilamenten, die als Glia-Zwischenfilamente bekannt sind und sich strukturell von Neurofilamenten dadurch unterscheiden, dass sie aus einem einzelnen 51 kDa-Protein bestehen und unterschiedliche physikalisch-chemische Eigenschaften aufweisen..
Alle Schichten, die Teil des Nukleoskeletts sind, sind tatsächlich Zwischenfilamentproteine. Diese haben ein Molekulargewicht zwischen 60 und 75 kDa und befinden sich in den Kernen aller eukaryotischen Zellen.
Sie sind wesentlich für die interne Organisation der Kernregionen und für viele Funktionen dieser Organelle, die für die Existenz von Eukaryoten wesentlich sind..
Diese Art von Zwischenfilament wiegt etwa 200 kDa und kommt überwiegend in Stammzellen des Zentralnervensystems vor. Sie werden während der neuronalen Entwicklung ausgedrückt.
Es gibt mehrere Krankheiten beim Menschen, die mit Zwischenfilamenten zusammenhängen.
Bei einigen Krebsarten wie beispielsweise malignen Melanomen oder Brustkarzinomen führt die Koexpression von Zwischenfilamenten von Vimentin und Keratin zur Differenzierung oder gegenseitigen Umwandlung von Epithel- und Mesenchymzellen.
Es wurde experimentell gezeigt, dass dieses Phänomen die wandernde und invasive Aktivität von Krebszellen erhöht, was wichtige Auswirkungen auf die für diesen Zustand charakteristischen metastatischen Prozesse hat..
Eriksson et al. (2009) untersuchen die verschiedenen Arten von Krankheiten und ihre Beziehung zu spezifischen Mutationen in den Genen, die an der Bildung der sechs Arten von Zwischenfilamenten beteiligt sind..
Krankheiten, die mit Mutationen in den Genen zusammenhängen, die für die beiden Keratinarten kodieren, sind Epidermolysis bullosa, epidermolytische Hyperkeratose, Hornhautdystrophie, Keratodermie und viele andere..
Zwischenfilamente vom Typ III sind an zahlreichen Kardiomyopathien und an verschiedenen Muskelerkrankungen beteiligt, die hauptsächlich mit Dystrophien zusammenhängen. Darüber hinaus sind sie auch für dominante Katarakte und einige Arten von Sklerose verantwortlich.
Viele neurologische Syndrome und Störungen sind mit Filamenten des Typs IV verbunden, wie z. B. Parkinson. In ähnlicher Weise sind genetische Defekte in Filamenten vom Typ V und VI für die Entwicklung verschiedener autosomaler Erkrankungen verantwortlich und hängen mit der Funktion des Zellkerns zusammen..
Beispiele hierfür sind unter anderem das Hutchinson-Gilford-Progerie-Syndrom, die Emery-Dreifuss-Muskeldystrophie..



Bisher hat noch niemand einen Kommentar zu diesem Artikel abgegeben.